АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
Общая характеристика эволюции генетического аппарата
Эволюционные возможности эукариотического типа клеточной организации превосходят таковые у прокариот. Указанную особенность объясняют, прежде всего, увеличением размеров ядерного генома. Количество генов в геноме бактерии и клетки человека соотносится как 1:100-1000. Известное значение могли иметь наличие в клетке двух аллелей каждого гена (диплоидность), многократное повторение некоторых генов. Все это расширяет масштабы мутационной изменчивости, совместимой с выживанием, и способствует образованию резерва наследственной изменчивости.
При переходе к эукариотическому типу усложнился также механизм регуляции жизнедеятельности клетки. На уровне генетического материала это проявилось в увеличении относительного числа регуляторных генов, замене единичных «голых» молекул ДНК множественными хромосомами, в которых ДНК образует комплекс с белками. В итоге оказалось возможным считывать информацию по частям с разных блоков генов в разных клетках и в разное время. В бактериальной клетке, например, одновременно транскрибируется 80-100 % информации генома. В клетках человека в зависимости от органа считывается 8-10 % (печень, почка, селезенка) до 44 % (головной мозг). Использованию информации по частям принадлежит исключительная роль в образовании многоклеточных живых форм. Определенное значение для перехода к многоклеточности имело, по-видимому, и приобретение клетками эукариотического типа эластичной оболочки, что позволяет образовывать стабильные комплексы клеток. В условиях усложнения генетического аппарата эукариот важным эволюционным приобретением стал митоз как механизм воспроизведения в поколениях генетически сходных клеток. Возникший путем эволюции митоза мейотический процесс интенсифицировал комбинативную изменчивость.
КЛЕТОЧНЫЙ ЦИКЛ
| Интерфаза (фаза между делениями клетки) | Пресинтетический период (G1) – 2n2c. Длительность ее непостоянна – от нескольких часов до нескольких суток. Дочерние клетки начинают свой цикл. Клетка содержит диплоидный набор хромосом, каждая хромосома содержит одну молекулу ДНК (2n2c). Наблюдаются интенсивные процессы биосинтеза, контролируемые ферментами. Клетка синтезирует структурные и функциональные белки, в том числе ферменты синтеза РНК и ферменты, необходимые для репликации ДНК. Ядрышко продуцирует рРНК, мРНК, тРНК; формирование рибосом. Образование эндоплазматической сети, лизосом, аппарата Гольджи, вакуолей и пузырьков, деление митохондрий и хлоропластов (у растений). Синтез АТФ в качестве запаса энергии, необходимой для процессов репликации и работы митотического аппарата. Синтез пуриновых и пиримидиновых оснований, входящих в состав ДНК. Образование веществ, подавляющих или стимулирующих начало следующей фазы. Если продуцируется фактор роста, то клетка переходит в S-период. Синтетический период (S) – 2n4c. Длительность фазы 6-8 часов. В различных типах клеток она меняется незначительно. Продолжают протекать биосинтетические процессы, начатые в G1-периоде. Синтез ДНК инициируется появлением в цитоплазме клетки сигнального фактора – активатора S-фазы. Для построения хроматина с высокой скоростью синтезируются гистоны в цитоплазме, которые затем мигрируют в ядро. Синтез ДНК – репликация, самоудвоение хромосом. Репликация центриолей (в тех клетках, где они имеются) и начало образования веретена деления. Построение второй хроматиды, в которую переходит вновь образовавшаяся молекула ДНК: получаются двухроматидные хромосомы. Постсинтетический период (G2) – 2n4c. Длительность фазы составляет 2-6 часов. Интенсивные процессы биосинтеза. Наблюдается активность ферментов, репарирующих повреждения в структуре ДНК. Происходит синтез РНК, синтез белков, необходимых для деления клетки, например, тубулина. Деление митохондрий и хлоропластов. Увеличение и запасание энергетических запасов. Масса цитоплазмы удваивается, возрастает объем ядра. |
 Морфологический критерий интерфазы: ядро в клетке округлое, с четкими границами. В нем видны 1 или 2 ядрышка. Хроматин в виде глыбок заполняет кариоплазму. Морфологический критерий интерфазы: ядро в клетке округлое, с четкими границами. В нем видны 1 или 2 ядрышка. Хроматин в виде глыбок заполняет кариоплазму.
| ||
| Фазы митоза | Профаза (первая фаза деления) – 2n4c Обычно самая продолжительная фаза клеточного деления. Появление в ядрах нитчатых структур – митотических хромосом (двухроматидные хромосомы). Хроматиды укорачиваются (до 4 % своей первоначальной длины) и утолщаются в результате их спирализации и конденсации. Этому событию предшествует повышение активности фосфорилаз, модифицирующих гистоны, и в первую очередь гистон Н1. В профазных хромосомах уже можно наблюдать зрелые кинетохоры, которые не имеют никаких связей с микротрубочками. Конденсация хромосом совпадает с резким уменьшением транскрипционной активности хроматина, которая полностью исчезает к середине профазы. В связи с падением синтеза РНК и конденсацией хроматина происходит инактивация и ядрышковых генов. Ядрышки исчезают, большая часть ядрышковых белков диссоциирует и в свободном виде встречается в плазме клетки или связывается с поверхностью хромосом. Происходит фосфорилирование белков ламины, ядерной оболочки, при этом теряется ее связь с хромосомами. Затем ядерная оболочка фрагментируется на мелкие вакуоли, а поровые комплексы исчезают. Собираются нити веретена деления. Активируются клеточные центры. В начале профазы разбираются микротрубочки в цитоплазме и начинается бурный рост множества астральных микротрубочек вокруг каждой из удвоившихся диплосом. Все микротрубочки, отходящие от центросом, растут вперед плюс-концами. В животных клетках и у низших растений центриоли расходятся к противоположным полюсам клетки. От каждой центриоли в виде лучей расходятся короткие микротрубочки, образующие в совокупности звезду. Наблюдается дезорганизация эндоплазматического ретикулума (он распадается на мелкие вакуоли, лежащие по периферии клетки) и аппарата Гольджи, который теряет свою околоядерную локализацию, распадается на отдельные диктиосомы, беспорядочно разбросанные в цитоплазме. | Морфологический критерий профазы:
Ядро заметно увеличивается, в нем исчезают ядрышки. В кариоплазме наблюдается образование, напоминающее клубок, составленный из тонких нитей. Эти тонкие нитевидные структуры – хромосомы. В конце профазы оболочка ядра разрушается и хромосомы выходят в цитоплазму.
|

Рис.
Профаза.
1-центросомы с парой центриолей; 2-веретено деления; 3-хромосомы;
4-центромеры; 5 фрагменты ядерной мембраны; 6- микротрубочки веретена деления; 7 микротрубочки, не связанные с кинетохором; 8 - микротрубочки, связванные с кинетохором.
|

Рис. Метафаза.
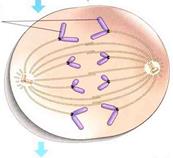 |
Рис. Анафаза.
1- сестринские хроматиды, ставшие дочерними хромосомами.
|
|
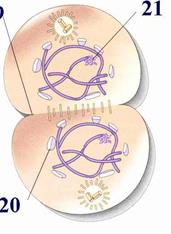
Рис. Телофаза.
1-образующееся
ядрышко;
2-формирующаяся клеточная перегородка;
3-восстанавливающаяся ядерная оболочка.