АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
ЦЧС- цитоплазматична чоловіча стерильність
Сукупність чоловічих генеративних органів квітки називають андроцеєм (від грецьк. «андрос» - чоловік). Формування мікроспор – мікроспорогенез - відбувається в пиляках тичинок – мікроспорангіях. Сформовані мікроспори проходять потім наступний цикл розвитку – гаметогенез, в результаті якого у квітці формується чоловічий гаметофіт.

Рис. Поступальні стадії формування пиляка пшениці (за Батигіна, 1987).
1-8- премейотичний період; 9-12-мейотичний період; 13-18-постмейотичний період. 1-8- премейотичний період; 9-12-мейотичний період; 13-18-постмейотичний період.
А- археспорій, ВСл – внутрішній шар, НСл - зовнішній шар, ПСл – парієтальний шар, СпТк – спорогенна тканина, СрСл- середній шар, Тп – тапетум, Эн - ендотецій, Эп – епідерма (екзотецій).
3.1. Пиляк і тичинкова нитка.

Рис. Розвиток пиляка: А-тичинки пасльону чорного. 1- пиляк, 2- тичинкова нитка; Б- почергові стадії розвитку пиляка (1-4), закладка первинного археспорію; Т – тапетум, СК-спорогенні клітини; В - окремий сектор із пиляка: 1- епідерміс, 2- ендотецій, або фіброзний шар, 3- середній шар, 4-тапетум, 5-материнські клітини мікроспор; Г- поперечний розріз пиляка евкаліпта: 1- одноядерний пилок, 2- зрілий пилок.
Тичинка покритонасінних рослин гомологічна мікроспорофілу, пиляк – мікроспорангію різноспорових плауноподібних і папоротеподібних, лусці і пиляку голонасінних. Згідно АВС- моделі закладка зачатків тичинок контролюється сумісно генами
групи В і С. Тичинки закладаються у вигляді горбиків (бугорків) слідом за горбиками оцвітини, на ранніх фазах розвитку квітки. В цей період структура пиляка мало чим відрізняється від структури тичинкової нитки і складається із паренхімних клітин меристематичної тканини. Пізніше, по мірі диференціації, пиляк набуває структури, яка різко відрізняється від структури тичинкової нитки. Відрізняються вони в першу чергу особливостями ділення ініціальних клітин. Наприклад, тичинкова нитка утворюється за рахунок апікального, а потім інтеркалярного ділення і росту клітин примордію. В дистальній частині ініціальні клітини започатковують тканини пиляка.
Тичинкова нитка розвивається із меристеми і складається із епідерми і рихлої паренхіми, всередині її міститься судино-волокнистий пучок із кільчастих і спіральних судин; елементи флоеми виражені нечітко. Нитки тичинок можуть зростатись або лише основами, або на всій протяжності, з утворенням тичинкового каналу (мальвові); у деяких родин, навпаки, пиляки зрощуються в пилкову трубку, а тичинкові нитки залишаються вільними(складноцвіті). Інколи тичинкова нитка не виростає зовсім або дуже коротка (сидячі пиляки), чи, навпаки, вона занадто довга і еластична (у вітрозапильних рослин - кукурудзи, жита - пиляки гойдаються). Тичинкові нитки цих рослин ростуть надзвичайно швидко. Мікроскопічні і гістохімічні дослідження росту тичинкових ниток кукурудзи показали, що клітини епідерми і паренхіми містять багато гетероауксину, під час росту ділення клітин не відбувається, а ріст відбувається за рахунок швидкого витягування клітин у поздовжньому напрямку.
Пиляк.
Кожний пиляк складається з двох пилкових мішків, сполучених в’язальцем. У двох ділянках кожного пилкового мішка під епідермою закладаються гнізда пиляка (мікроспорангіїї). Клітини спорангію великі з густою цитоплазмою, крупними ядрами і ядерцями. Їх називають первинні клітини археспорію. У різних рослин число клітин археспорію різне: від 1-3 до десятків і навіть сотень тисяч, як це, наприклад, у видів Triticum, різних родів орхідей. Відповідно з цим і число пилкових зерен в пиляках різних рослин різне: від декількох до чотирьох мільйонів, як це у видів Саttleya. Комплекси цих клітин називаються спорогенною тканиною або археоспорій пиляка. (рис.13.Устинова с 73).
Надалі первинні клітини археспорію діляться і утворюють парієнтальний шар клітин (прилягає до епідерми) і вторинні клітини археспорію, що перетворюються у материнську клітину мікроспор,або мікроспороцити.
Ми спочатку розглянемо формування і будову стінки пиляка, а потім онтогенез материнських клітин.
Пиляк зверху покривають клітини епідерми. Зовнішні стінки оболонки епідермальних клітин кутинізовані, потовщені, інколи покриті воском. Клітини епідерми пиляків містять хромопласти.
Під епідермою у більшості покритонасінних рослин утворюється особливий - фіброзний (волокнистий) шар. Його клітини містять хлоропласти. Стінки клітин фіброзного шару утворюють різноманітні потовщення, які сприяють розтріскуванню пиляка. Проте не всі пиляки тріскаються таким чином. У деяких рослин пиляки розкриваються з допомогою особливих пор, дірочок і клапанів. Пиляки ж клейстогамних рослин фіброзного шару не утворюють – пилок у них проростає в пиляках. Фіброзні потовщення відсутні також у водних рослин. У зв’язку з тим, що фіброзні потовщення виражені не завжди, то шар клітин, що прилягає до епідерми називають ендотецієм, на відміну від екзотеція, до якого відносять епідерму.
Глибше під фіброзним шаром розміщується шар клітин, який називають тапетум.
Клітини тапетуму на ранніх етапах розвитку одноядерні, пізніше стають дво- і навіть багатоядерними.
Походження тапетуму може бути подвійним: він розвивається із найглибшого внутрішнього шару клітин, або ж розвивається із клітин первинного археспорію.
Тапетум починає лізуватись після мейозу в материнських клітинах на стадії розпаду
тетрад мікроспор і у зрілому пиляку його немає.
Тапетум пиляка має важливе значення у формуванні пилку, бо усі пластичні речовини
для живлення спорогенної тканини і материнських клітин мікроспор надходять саме через нього.
Ядра в клітинах тапетуму діляться мітотично за трьома типами:
1) нормальний мітоз без цитокінезу – ядра зближені, ділення відбувається звичним чином. Клітини двоядерні, рідше чотириядерні. Цей тип поділу клітин тапетуму у рослинному світі найбільш поширений.
2) відстаючий тип ділення – помилкове розходження хромосом в анафазі і утворення містків із відстаючих хромосом, що спричинює утворення гантелеподібних ядер. Описано цей тип ділення вперше у лілейних Купером (1933).
3) ендомітоз – ділення в середині ядра зі збереженням ядерної оболонки і ядерця.
Всі утворені хромосоми залишаються в ядрі і клітини тапетуму містять тетраплоїдні ядра. Цей тип ділення вперше описаний у складноцвітих і пасльонових.
За будовою розрізняють два види тапетуму:
1) Секреторний, або залозистий, - клітини зберігають своє положення і виділяють особливу рідину – секрет, оболонки клітин не розчиняються, ядра гіпертрофовані.
Амебоїдний, або периплазмодій - стінки клітин тапетуму частково щезають і їхні протопласти проникають між материнськими клітинами мікроспор (нетиповий
периплазмодій). Відомий і інший вид периплазмодію, коли протопласти клітин зливаються в загальну масу і виникає типовий периплазмодій.
Не зважаючи на наявність в пиляку морфологічно і біохімічно різних тканин, усі процеси в ньому, як і в інших генеративних органах, відбуваються чітко і впорядковано. Всі тканини пиляка зв’язані між собою плазмодесмами, що полегшує надходження поживних речовин до мікроспороцитів. Тільки під час мейозу, коли мікроспороцити діляться і оточені калозною оболонкою, цей зв’язок порушується.
3.2. Мікроспорогенез – це процес утворення мікроспор із материнських клітин.
Під час мікроспорогенезу в материнських клітинах відбувається два поділи
(редукційний і екваційний) в результаті утворюються тетради мікроспор, які надалі розвиваються у чоловічі гамети. Пригадаємо, що саме мейоз забезпечує перехід від диплоїдного стану до гаплоїдного і відбувається розділ материнської клітини пилку на чотири клітини.

Рис. Сукцесивний тип розвитку тетрад мікроспор.
1-7 – мейоз; 7- ізобілатеральне розміщення мікроспороцитів; 8-9- мікроспори; Д-діада;
К-калоза; ТМ-тетрада мікроспор.
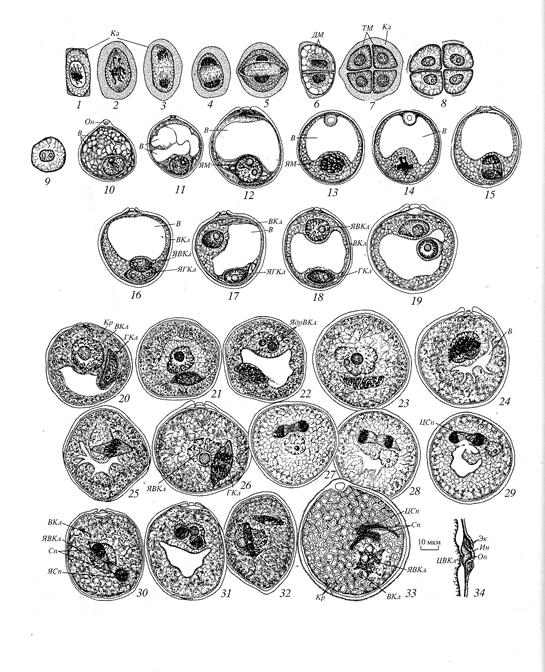
Рис. Мікроспорогенез і розвиток пилкового зерна у пшениці (за Батыгина, 1987)
В-вакуоля; ВКл – вегетативна клітина; ГКл –генеративна клітина; Дм – діада;
И- інтина; К - калоза; Кр – крохмаль; Оп – оперкулюм (кришечка); П - пора; Сп - спермій; ТМ - тетрада мікроспор; ЦВКл - цитоплазма вегетативної клітини; ЦСп – цитоплазма спермію; Э – екзина; ЯВКл – ядро вегетативної клітини; ЯМ – ядро мікроспороциту; ЯСп – ядра сперміїв; ЯдВКл – ядерце вегетативної клітини.
В одному й тому ж пиляку мейоз відбувається або синхронно, і у такому разі усі материнські клітини мікроспор перебувають на однаковій фазі ділення, або асинхронно, і тоді в одному й тому ж пиляку матимуть місце різні фази ділення. Асинхронність мейозу особливо добре проявляється у суцвіттях, де можна спостерігати усі фази мейозу, від початку його виникнення до повного його завершення.
Перетворення материнських клітин в тетради мікроспор у різних видів проходить по різному. Встановлено три типи утворення тетрад мікроспор: сукцесивний (послідовний), проміжний і симультанний (одночасний).
При сукцесивному розвитку після першого ділення мейозу утворюється перетинка із фрагмопласта веретена, яка поділяє материнську клітину на дві дочірні і утворюється діада клітин. Після другого поділу мейозу знову закладається перегородка в кожній із дочірніх клітин, в результаті протопласт материнської клітини послідовно розділяється на чотири частини. Такий тип утворення тетрад властивий переважно однодольним і лише зрідка зустрічається у деяких видів дводольних.
За цього типу розвитку тетрад часто має місце інтерфаза між першим і другим діленням мейозу. Залежно від умов погоди під час мікроспорогенезу інтерфаза може бути довготривалою.
У тому разі, коли після першого ділення мейозу клітинні перетинки не утворюються і усі чотири клітини виникають одночасно після другого поділу завдяки тому, що на периферії клітини виникають чотири борозенки, які розділяють одночасно від периферії до центру вміст клітини на 4 частини. Отже, при цьому типі утворення тетрад завжди протопласт материнської клітини одночасно ділиться на 4 клітини – мікроспори.
Сукцесивний тип розвитку тетрад звично має місце у однодольних, симультанний – у дводольних, бувають виключення.
Деяким видам (магнолієвим) притаманний третій тип утворення тетрад – проміжний:
тут після першого ділення мейозу перетинка між ядрами не утворюється, а цитоплазма починає перешнуровуватись на дочірні клітини, що завершується після другого ділення мейозу,
Більшість ембріологів вважають симультанний тип більш примітивним, а сукцесивний – більш прогресивним.
Описано 5 типів розміщення мікроспор в тетрадах:
- тетраедральне,
- ізобілатеральне,
- хрест – нахрест,
- Т- подібне,
- лінійне:

Рис. Схема різних типів розміщення мікроспор в тетрадах:
А- тетраедральне; Б - ізобілатеральне; В – хрест-нахрест; Г- Т-подібне; Д - лінійне.
При сукцесивному способі розвитку мікроспори розміщуються у переважної більшості видів ізобілатерально, при симультанному – у формі тетрад. Лінійне розміщення зустрічається дуже рідко
Тривалість перебування мікроспор у стані тетрад у більшості однорічних рослин, що цвітуть у жаркий час, складає декілька годин, рідше одну добу (у традесканції 22 год).
Поиск по сайту: