АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
Комплементарность
|
Читайте также: |
У популярного генетического объекта плодовой мушки Drosophila melanogaster имеется большое число форм, наследственно различающихся по окраске глаз. У мух так называемого дикого типа или типа, распространенного в природе, глаза темно-красные. Существуют формы с ярко-красными глазами. Этот признак рецессивен по отношению к дикому типу. Он наследуется по моногибридной схеме при скрещивании нормальных мух и мух с ярко-красными глазами. Соответствующий ген обозначается: st (scarlet) - рецессив; st+ - доминант.
Существуют также мухи с коричневыми глазами. Это тоже рецессивный признак, наследующийся по моногибридной схеме при скрещивании диких мух и мух с коричневыми глазами. Соответствующий ген обозначается bw (brown) - рецессив; bw+ - доминант1.
Если скрестить мух с ярко-красными глазами и мух с коричневыми глазами, то получаются следующие результаты. В Fi все мухи имеют темно-красные глаза (дикий тип), а при скрещивании гибридов первого поколения в F2 появляются четыре класса расщепления: мухи с темно-красными, ярко-красными, коричневыми и белыми глазами в соотношении 9:3:3:1. Для объяснения этого результата обратимся к логике генетического анализа.
Результаты, получившиеся в F1, показывают, что существует некоторый тип взаимодействия. Можно предположить, что это взаимодействие аллелей одного гена при моногибридном скрещивании. Однако в F2 появляются четыре класса в соотношении, характерном для дигибридного скрещивания при полном доминировании по обоим признакам, и среди них формируется самый малочисленный класс - мухи с белыми глазами.
Если справедливо предположение о том, что это скрещивание дигибридное, то генотипы родительских форм записывают:

где знак «+» соответствует нормальным (доминантным) аллелям генов: bw и st. Тогда самки и самцы образуют по одному типугамет - соответственно st bw+ и st+bw, а генотип гибридов F1 будет: st+st bw+bw.
Такие дигетерозиготные мухи должны образовать четыре типа гамет, которые во всевозможных сочетаниях при скрещивании между собой гибридов F1 дадут в F2 следующее расщепление по генотипу.
С помощью фенотипических радикалов можно написать следующее расщепление по фенотипу:
9 st+ - bw+ - - с темно-красными глазами
3 st+ - bw bw - с коричневыми глазами
3 st st bw+ - - с ярко-красными глазами
1 st st bw bw - с белыми глазами
Учитывая, что аллели st и bw рецессивны, можно объяснить появление первых трех фенотипических классов при расщеплении. При наличии нормальных аллелей st+ и bw+ мухи должны принадлежать к дикому типу по окраске глаз (9 с темно-красными глазами). При гомозиготности только по рецессивной аллели bw bw мухи должны быть с коричневыми глазами (3), так же как при гомозиготности только по st st мухи должны быть с ярко-красными глазами (3). Наконец, остается последний класс - двойной гомозиготный рецессив (1 st st bw bw), который соответствует мухам с белыми глазами. Все эти выводы можно проверить, исследуя далее расщепление при анализирующих скрещиваниях и скрещиваниях между особями F1.
Таким образом, предположение о дигибридном расщеплении в рассмотренном скрещивании подтверждается, а новообразование - белоглазые мухи в F2 - результат взаимодействия рецессивных аллелей st и bw.
В рассмотренной схеме также наблюдалось взаимодействие генов в F2, в результате которого дрозофилы имели нормальный цвет глаз. Такой тип взаимодействия носит название комплементарности или комплементарного (взаимно дополнительного) действия, когда доминантные аллели обоих генов обусловили нормальный (или дикий) фенотип (под комплементарностью обычно подразумевают именно этот тип взаимодействия генов).
В F2, рецессивные аллели тех же генов обусловили появление белоглазых мух. Был приведен пример того, что носит название формально-генетического анализа, при котором полностью абстрагируются от механизмов действия исследуемых генов. Если анализ проведен верно и формальные отношения генов и аллелей выявлены правильно, то последующее выяснение физиологических механизмов, лежащих в основе генных взаимодействий, только подтверждает и конкретизирует выводы.
Биохимический механизм взаимодействия аллелей генов st и bw исследован достаточно подробно. Известно, что у дрозофилы окраска глаз обусловлена синтезом двух пигментов - красного и бурого. Рецессивная аллель bw в гомозиготе прерывает синтез красного пигмента, поэтому глаза содержат только бурый пигмент. Рецессивная аллель st в гомозиготе блокирует синтез бурого пигмента, вследствие чего в глазах мух содержится только красный пигмент.
Когда в дигетерозиготе оказываются нормальные аллели обоих генов, синтезируются оба пигмента. Результат - комплементарное взаимодействие нормальных аллелей, наблюдаемое в F1. Если в F2 в гомозиготе оказываются и bwbw, и stst, то не синтезируются ни красный, ни бурый пигменты, и глаза оказываются белыми, неокрашенными.

Рис. 2 - Комплементарное взаимодействие генов, определяющих форму гребня у кур (соотношение 9:3:3:1)
Можно рассмотреть и другой тип комплементарного взаимодействия генов у дрозофилы, если идти от метаболического эффекта генов к схеме их взаимодействия. Вновь обратимся к биосинтезу пигментов глаза у дорозофилы (рис. 3). Известно, что кроме рецессивных аллелей st синтез бурого пигмента блокируют и рецессивные аллели гена purple (pr). Фенотип гомозигот stst и ргрт - ярко-красные глаза. При скрещивании таких мух в F1 глаза нормальные - темно-красные, поскольку работают оба гена - комплементарно взаимодействуют их доминантные аллели. В F2 наблюдается следующее соотношение фенотипов: 9 с темно-красными и 7 с ярко-красными глазами. Это объясняется тем, что выход в гомозиготу любой из двух рецессивных аллелей достаточен для блокирования синтеза пигмента, тем более, когда и рг, и st находятся в гомозиготе. Это тоже пример комплементарного взаимодействия, но без новообразования в F2.
По типу комплементарное™ взаимодействуют гены, контролирующие разные этапы одного и того же метаболического пути. Однако для многих морфологических признаков неизвестен биохимический механизм их реализации, поэтому приходится ограничиваться констатацией формально-генетической схемы их наследования. Так, по типу комплементарности взаимодействуют гены, определяющие форму гребня кур (рис. 2), форму плода у тыквы (рис. 3) и др.
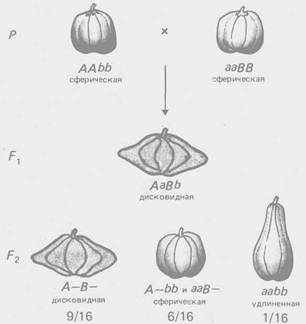
Рис. 3 - Комплементарное взаимодействие генов, определяющих форму плода тыквы (соотношение 9:6: 1)
Эпистаз
Вернемся к анализу взаимодействия генов рr и st у дрозофилы. Соотношение фенотипических классов в F2 можно представить себе и как следствие того, что рецессивная аллель рr в гомозиготе препятствует проявлению доминантной аллели st+. Точно так же рецессивная аллель st в гомозиготе препятствует проявлению доминантной аллели рr+. Действительно, то, что известно о генетическом контроле синтеза бурого глазного пигмента у дрозофилы, вполне соответствует предложенному здесь объяснению. Такой тип взаимодействия носит название эпистатического, или эпистаза, и условно изображается: рr > st+ и st > pr+. В данном случае рецессивная аллель рr эпистатична по отношению к доминантной аллели st+, a st эпистатична по отношению к рr+. Данный случай взаимодействия генов называют также двойным рецессивным эпистазом.
По изменению числа и соотношения классов дигибридного расщепления в F2рассматривают несколько типов эпистатических взаимодействий: простой рецессивный эпистаз (а > В; а> Ь или b > А; Ь > а), который выражается в расщеплении 9:3:4; простой доминаданый эпистаз (А> В; А> b или В > А; B > а) с расщеплением 12:3:1 и т.д.
Один ген, подавляющий действие другого, называют эпистатическим геном, ингибитором или супрессором. Подавляемый ген носит название гипостатического.
Как уже показано, констатация того или иного типа взаимодействия генов в дигибридном скрещивании условна. Тем не менее, при кажущемся нарушении закона независимого наследования (появлении неожиданных классов в расщеплении или уменьшении числа классов), связанного с взаимодействием двух генов, всегда можно свести наблюдаемые соотношения в F2 к классическому 9:3:3:1. При этом важно понять, какие классы объединились, и тогда интерпретировать тип взаимодействия.
Необходимо также отметить, что само словосочетание «взаимдействие генов» условно. В действительности взаимодействуют продукты генов, а не сами гены, так что правильнее было бы говорить о взаимодействии фенов, а не о взаимодействии генов. Отсюда понятно, что судить о том, с каким скрещиванием имеет дело экспериментатор: моногибридным, дигибридным или полигибридным - можно только на основании результатов полного гибридологического анализа.
Полимерия
Наряду с комплементарным и эпистатическим принято также рассматривать взаимодействие генов по типу полимерии. В этом случае разные гены как бы дублируют действие друг друга, и одной доминантной аллели любого из взаимодействующих генов достаточно для проявления изучаемой фенотипической характеристики. Так, при скрещивании растений пастушьей сумки с треугольными плодами (стручками) и с овальными плодами в F1 образуются растения с плодами треугольной формы. При их самоопылении в F2наблюдается расщепление на растения с треугольными и овальными стручками в соотношении 15:1. Это объясняется тем, что существуют два гена, действующих эднозначно. В этих случаях их обозначают одинаково (А1 и А2). Тогда все генотипы: А1 - А2 -, А1 - а2а2, а2а2А2 - будут иметь одинаковую фенотипическую характеристику - треугольные стручки, и только растения а1а1а2а2 будут отличаться - образовывать овальные стручки. Это случай так называемый некумулятивной полимерии.
Однозначные, или полимерные, гены могут действовать и по типу кумулятивной полимерии. Так, шведский генетик Г. Нильсон-Эле в 1908 г. описал серию однозначно действующих генов, которые определяют окраску эндосперма зерен пшеницы. При этом интенсивность окраски зерен оказалась пропорциональной числу доминантных аллелей разных генов в тригибридном скрещивании. Наиболее окрашенными были зерна А1А1А2А2А3А3, а зерна а1а1а2а2а3а3 не имели пигмента. Между этими крайними типами при расщеплении в F2 наблюдались промежуточные варианты в соотношении 1:6:15:20:15:6:1.
По типу кумулятивной полимерии наследуются многие количественные признаки, например цвет кожи у человека; молочность, яйценоскость, масса и другие признаки сельскохозяйственных животных; длина колоса у злаков, содержание сахара в корнеплодах сахарной свеклы и др. Изучением наследования таких признаков занимается специальный раздел генетики - генетика количественных признаков, которая важна прежде всего для селекции и разработки проблем микроэволюции.
6. Влияние факторов внешней среды на действие генов
Влияние внешней среды: условий питания, температуры, света, химического состава и структуры почвы, влажности и т.п. - на индивидуальное развитие организма огромно. Человеку, занимающемуся сельскохозяйственным производством, очевидна роль внешней среды в формировании организмов. Внешняя среда определяет процессы отбора, изменчивости и наследственности, т.е. факторы эволюции. В соответствии с изменениями внешней среды развивался весь органический мир. Благодаря этому каждый вид организма приобрел своеобразное внутреннее строение и присущий ему тип индивидуального развития, закрепленные наследственностью.
Поскольку у организмов разных видов благодаря совместному действию указанных трех факторов эволюции выработались приспособления к определенным условиям внешней среды, их нормальное развитие стало возможно только при данных условиях. Изменение последних может приводить к изменению наследственных факторов (мутации), с чем мы познакомимся позднее, и к изменению процессов индивидуального развития, вызывающему видоизменение фенотипа, т.е. к изменению проявления действии генов. Реализация наследственного признака или свойства организма является результатом взаимодействия генотипа и условий внешней среды.
Создавая определенные внешние условия, можно направлять действие генов в нужную нам сторону, т.е. управлять процессами индивидуального развития организма. Генетики давно обратили на это внимание. Проблема управления индивидуальным развитием привлекла внимание И.В. Мичурина. Он показал, что наследственная потенция организма огромна, и, подставляя соответствующие внешние условия для развития гибридов плодовых и ягодных растений, можно изменять доминирование и выявлять наиболее ценные наследственные качества и свойства организма.
По этому поводу И.В. Мичурин в 1929 г. писал: «В общем влияние всей совокупности внешних факторов на строение организма гибридов настолько велико, что в большинстве случаев значительно подчиняет себе действия наследственной передачи качеств и свойств растений-производителей. В особенности такое влияние резко отражается на состоянии материнского растения при закладке у него в строении семени зачатков будущего организма гибрида и на полученном гибриде в самой ранней стадии его развития, благоприятствуя одним и являясь непреодолимым препятствием для проявления других наследственных признаков». При этом И. В. Мичурин добавлял, что «в законе Менделя нисколько не отвергаю его достоинств, напротив, я лишь настаиваю на необходимости внесения в него поправок и дополнений...».
Одним из ярких примеров влияния внешней среды на действие гена и выражение признака является изменение доминирования. Так, И.В. Мичурин на гибриде груши маслинолистной (Pyrus elaeagnifolia) и сорта Бессемянка и на гибриде вишен Самарской и Владимирской показал, что в гибридном организме будут доминировать те свойства и признаки, которые встречают наиболее благоприятные условия для своего осуществления. Для той же цели управления доминированием им применялся «спартанский режим воспитания» гибридных сеянцев, а также метод ментора.
Изучение влияния внешних факторов на действие отдельных генов у животных также показало возможность изменять фенотип организма. Так, например, встречаются кролики с желтым и белым жиром. Кролики с желтым жиром имеют в своем генотипе рецессивную аллель у в гомозиготном состоянии. Наличие доминантной аллели гена У определяет белый жир. Этот ген контролирует выработку энзима, который разрушает желтый пигмент - ксантофилл, содержащийся в зеленом корме. Если кроликам с желтым жиром (по генотипу уу) давать корм, который не содержит ксантофилла, то у них будет образовываться белый жир, так как для реализации действия гена не будет создано подходящих условий.
У гибридов от скрещивания озимых и яровых форм пшеницы или ржи в первом поколении доминирует свойство яровости. Во втором поколении у ржи наблюдается расщепление в отношении 3:1, у пшениц могут быть расщепления 3:1 и 15:1 и более сложные. Если семена второго поколения посеять весной, то озимые формы будут куститься, но не будут колоситься, что даст возможность обнаружить расщепление. Если же эти семена предварительно прояровизировать, т.е. подвергнуть предпосевной обработке низкими температурами, то все растения второго поколения будут колоситься в год посева. В этом случае мы не обнаружим расщепления.
У одноклеточной зеленой водоросли хлореллы известны наследственные изменения, обусловливающие отсутствие хлорофилла. Эти формы не могут жить и размножаться в обычных условиях на свету, так как они не способны осуществлять фотосинтез. Но если в питательную среду для таких форм добавить глюкозу, то они смогут размножаться. Подобные опыты проводились и с бесцветными альбинотическими проростками кукурузы.
Можно твердо сказать, что бессмысленно представлять себе проявление наследственности вне конкретных условий внешней среды. Знание механизмов действия гена и особенностей влияния внешней среды на его проявление обеспечивает возможность управления развитием врожденных свойств. Благодаря этому развитие наследственных заболеваний у человека перестает быть фатальной неизбежностью. Изучение влияния внешних условий на породные и сортовые (т.е. наследственные) хозяйственно ценные свойства организмов расширяет возможности повышения продуктивности животных, растений и микроорганизмов.
Выводы
Итак, подводя итоги нашей работы можно сказать, что генетический анализ наследования и взаимодействия отдельных генов у организмов независимо от их филогенетического уровня и сложности организации служит необходимым начальным приемом, соответствующим качественному и количественному анализу в химии. Когда исследователь имеет дело с неизвестным ему химическим веществом, он в определенной последовательности исследует сначала группы элементов, а затем сами элементы и, наконец, их количественное соотношение.
Однако следует иметь в виду, что сам по себе генетический анализ взаимодействия генов является формальным, так как наследственные признаки используются в нем лишь в качестве «маркеров»для прослеживания судьбы генов в ряду поколений и их взаимосвязи на «финише» индивидуального развития, т.е. в фенотипе. Взаимодействие генных продуктов при формировании признаков данному анализу оказывается недоступным.
В ходе изложения неоднократно обращалось внимание на то, что все перечисленные закономерности поведения генов в хромосомах оказываются общими для животных и человека, для высших и низших растений, а также для одноклеточных организмов.
Из анализа расщепления, независимого комбинирования генов, а также их взаимодействия видно, что комбинация генов и их взаимодействие создают огромную наследственную изменчивость. В результате комбинации генов могут возникать новые наследственные признаки. Такого типа изменчивость называется комбинативной изменчивостью. Она играет существенную роль эволюционном процессе как источник изменчивости, как механизм, обеспечивающий сочетание наиболее приспособительных признаков для существования организмов.
Поиск по сайту: