АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
ИНБРИДИНГ И ГЕТЕРОЗИС
|
Читайте также: |
Закон Харди-Вайнберга действует только тогда, когда скрещивание проходит случайно, т. е. когда вероятность скрещивания между двумя генотипами равна произведению их частот. Такая случайно, свободноскрещивающаяся популяция называется панмиктической. Она претерпевает панмиксис по отношению к рассматриваемым признакам, которые контролируются генотипом.
Когда скрещивание неслучайно, т. е. особи с определенными генотипами (сходными или различными) спариваются между собой чаще, чем этого следует ожидать на основе случайности, говорят об ассортивном скрещивании. Генетическим результатом здесь оказывается получение большего количества гомозигот, чем должно быть при свободном скрещивании.
Интересную форму ассортивного скрещивания представляет собой инбридинг, при котором скрещивания между родственными особями происходят чаще, чем можно было ожидать на основе случайности. Поскольку родственные особи в генетическом отношении более сходны между собой, чем не состоящие в родстве организмы, инбридинг ведет к повышению частоты гомозигот и снижению частоты гетерозигот по сравнению с теоретически ожидаемой при случайном скрещивании, частоты аллелей при этом не изменяются.
Существуют различные степени инбридинга. Самым крайним случаем инбридинга является самооплодотворение, или самоопыление. В мире животных таких организмов немного, а среди домашних животных их вообще нет. В мире растений самоопыляющихся видов много. Это пшеница, ячмень, овес, бобы, соя, томаты и др.
Применяют инбридинг для разложения популяций на гомозиготные линии, легче всего это удается у растений-самоопылителей. Представим себе гипотетический случай – самоопыляющееся растение, гетерозиготное по одной паре аллелей одного локуса. Условно примем, что каждое растение дает четырех потомков (табл. 14).
Анализ данных таблицы показывает, как быстро при данной ситуации возрастает гомозиготность. Доля гетерозигот в каждом из трех первых поколений уменьшается в два раза, а доля гомозигот, наоборот, возрастает в два раза. В дальнейших поколениях доля гетерозигот в каждом из них будет систематически уменьшаться вдвое, а через 10 лет будет достигнута почти полная гомозиготность. Потомство таких гомозиготных растений называют чистыми линиями.
Таблица 14
Влияние самоопыления на изменение доли гомозигот
в ряду поколений у растения, исходное поколение которого
было моногибридным
| Поколение | Соотношение генотипов | Доля гетерозигот | Доля гомозигот | ||
| АА | Аа | аа | |||
| F1 | |||||
| F2 | |||||
| F3 | |||||

| 
| 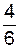
| |||
| или | |||||
| F4 | |||||
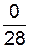
| 
| 
| |||
| или | 12,5 | 87,7 |
При других степенях инбридинга повышение гомозиготности будет протекать медленнее (брат × сестра, полубрат × полусестра, двоюродный брат с сестрой, отец × дочь, мать × сын и т. д.). Постоянное скрещивание полных братьев и сестер на протяжении десяти поколений приведет теоретически к созданию популяции, в которой 91,3% всех локусов будут гомозиготными. С. Райт высчитал, что скрещивание брат × сестра уменьшает остаточную гетерозиготность в каждом поколении на 19%, а при скрещивании полубрат × полусестра она уменьшается на 11%.
Все типы инбридинга повышают гомозиготность. Частота гомозигот в популяции возрастает за счет гетерозигот. Степень уменьшения гетерозиготности из-за инбридинга называют коэффициентом инбридинга F. Он представляет собой вероятность того, что у какой-либо особи в данном локусе окажутся два аллеля, идентичные по происхождению. В первом поколении самоопыления доля особей, несущих по два идентичных по происхождению аллеля, равна общей частоте гомозигот в популяции, то есть ½, и, следовательно, коэффициент инбридинга F = ½. В каждом последующем поколении инбридинга значение F возрастает на половину частоты гетерозигот в предыдущем поколении. Для определения доли гомозигот в поколениях инбридинга у растений используют формулу Райта: F = 1 – (1/2)n, где n – число поколений инбридинга.
Однако у видов, обычно перекрестноопыляющихся, инбридинг приводит к снижению жизнеспособности, которое непосредственно связано с гомозиготностью, сопровождается появлением летальных признаков или других дефектов. По мере того, как гены, ответственные за эти явления, переходят в гомозиготное состояние, они, конечно, элиминируют. Но элиминация нежелательных генов связана с потерей и многих желательных генов, потому что первые сцеплены со вторыми. Вот почему некоторые сильно инбридированные линии обладают невысокой продуктивностью и многие из них гибнут. У выживших при таком процессе очищения от нежелательных генов линий наблюдается явление, получившее название инбредной депрессии количественных признаков.
Итак, инбридинг увеличивает гомозиготность, а гомозиготность может привести к появлению дефектов; однако не следует делать вывод, что инбридинг не выгоден при любых обстоятельствах. В результате инбридинга многие особи становятся гомозиготными по вредным генам, но столько же особей становятся гомозиготными и по нормальным аллелям. Жизнеспособность самоопыляющихся видов показывает, что гомозиготность благоприятна.
У таких растений инбредная депрессия не возникает благодаря тому, что естественный отбор поддерживает в популяциях этих растений значительно более низкую частоту вредных рецессивных аллелей, чем в случайно скрещивающихся популяциях. В популяциях самоопылителей гомозиготность очень высока. В результате рецессивные вредные аллели элиминируют под действием естественного отбора по мере того, как они переходят в гомозиготное состояние.
Противопоставить инбредной депрессии можно скрещивание между разными независимыми инбредными линиями. У таких гибридов заметно возрастает приспособленность в отношении плодовитости, жизнеспособности, размеров и т. д. Это явление называется гибридной мощностью, или гетерозисом. Гетерозис проявляется в превосходстве гибрида над обеими родительскими формами.
Независимые инбредные линии обычно становятся гомозиготными по различным рецессивным аллелям. При скрещивании их в потомстве можно сохранить гомозиготность по искусственно отобранным признакам, тогда как вредные аллели переводятся в гетерозиготное состояние.
Гетерозис все еще представляет собой одну из самых больших загадок генетики. Существуют несколько различающихся между собой теоретических объяснений причин его возникновения. Одно из них названо теорией доминантных генов, сторонники которой считают, что гетерозис обусловлен действием доминантных генов, тогда как рецессивные гены подавляют его.
Поскольку жизнеспособность и продуктивность организма обусловлены эффектом многих генов, маловероятно, чтобы инбредные линии были гомозиготными по всем этим генам. Потомство, полученное при скрещивании двух линий, будет представлено всеми доминантными генами, по которым каждый из родителей был гомозиготным. Это и обуславливает гетерозис. Если две независимые линии обладают пятью локусами, то скрещивание их даст следующие результаты.
Родители … ………………… ААввссDDее х ааВВссddЕЕ
Число локусов с доминантными генами 2 × 2
Потомство …………………………………… АаВвссDdЕе
Число локусов с доминантными генами 4
В данном случае у гетерозиготы все рецессивные гены, кроме одного, маскируются доминантными аллелями. Если данная теория верна, у потомства должен проявиться гетерозис. Можно предположить, что доминантный ген у гетерозиготы защищает ее от вредного воздействия рецессивного аллеля. Это можно представить еще и так: АА = Аа > аа.
Вторая теория, объясняющая явление гетерозиса, получила название сверхдоминирования. Эта гипотеза объясняет случай, когда эффект пары аллелей в гетерозиготном состоянии часто отличается от эффекта, проявляемого каждым из них в гомозиготном состоянии, и гетерозигота превосходит любую из родительских, гомозиготных по одному или другому из этих аллелей, форм. Другими словами, гибридная мощность объясняется преимуществом гетерозиготного состояния (АА < Аа > аа), когда доминантный и рецессивный аллель как бы дополняют друг друга при действии на признак. При этом эффект сверхдоминирования в гетерозиготе может наблюдаться даже в том случае, когда рецессивный аллель в гомозиготе летален или приводит к снижению жизнеспособности.
Трудность, с которой сталкиваются сторонники этой теории, заключается в том, что таких примеров, где гетерозиготность по одному локусу вызвала бы превосходство гибрида над его обоими родителями, не так много.
Однако ни одна из этих гипотез не может объяснить всех случаев гетерозиса. В.А. Струнников предложил теорию компенсаторных факторов. Этот термин обозначает комплекс генов и их аллелей, который подбирается при получении инбредных линий. Компенсаторный комплекс генов нивелирует отрицательные эффекты высокого уровня гомозиготности, а при гибридизации дает гетерозис.
Поскольку гетерозис не может касаться одновременно всех признаков растения или животного, его классифицируют на несколько типов.
1. Репродуктивный гетерозис, который выражается в лучшем развитии органов размножения, приводит к повышению урожайности плодов и семян.
2. Соматический гетерозис, приводящий к мощному развитию вегетативной массы.
3. Приспособительный, или адаптивный, гетерозис, который выражается в общем повышении жизнеспособности. Выделение этого типа гетерозиса связано с тем, что мощное развитие какого-либо признака не означает повышение адаптивной ценности организма. Например, крупнозерные сорта злаков обычно менее засухоустойчивы, увеличение длины стебля ведет к полеганию растений и т.д.
На проявление гетерозиса оказывает влияние направление скрещивания, условия выращивания гибридов F1, а также комбинационная способность линии, то есть способность образовывать продуктивные гибриды.
Большое внимание в генетических исследованиях уделяют изучению степени и характера проявления гетерозиса у гибридов первого поколения. Их оценивают по элементам продуктивности и устанавливают степень проявления истинного, гипотетического и конкурсного гетерозиса.
Истинный гетерозис (Гист) – способность растений F1 превосходить по конкретному признаку или их комплексу лучшую из родительских форм. Его определяют в процентах по следующей формуле:
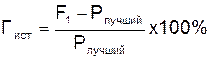 .
.
Гипотетический гетерозис определяется в процентах по отношению к средней выраженности данного признака у родительских форм (Рср). Вычисляют по формуле
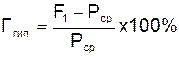 .
.
Однако истинный и гипотетический гетерозис не характеризуют ценность данной комбинации, ее характеризует конкурсный гетерозис (Гконк). Он показывает, на сколько процентов растения F1 данной гибридной комбинации превосходят районированный сорт или гибрид. Конкурсный гетерозис рассчитывают по формуле
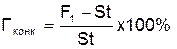 .
.
Во втором поколении гетерозис снижается, и это можно учесть:
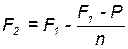 ,
,
где Р - средняя урожайность родительских линий, n – число линий, F1 – фактическая урожайность гибридов первого поколения, F2 – предполагаемая урожайность F2.
Задачи
1. Дайте точное определение различий между: инбридингом, инцухтом, инбредной линией, инцухт-линией и чистой линией.
2. Где следует ожидать большей доли летальных генов среди их общего числа – у фасоли или у кукурузы и почему?
3. Следует ли считать инбридинг и ассортивное скрещивание взаимоисключающими друг друга отклонениями от свободного скрещивания (панмиксиса)? Объясните.
4. В двух инбредных линиях кукурузы и у их гибридов (F1) исследовали активность поглощения кислорода. У гибридов активность поглощения кислорода оказалась выше, чем у родительских линий. Предложите генетическую гипотезу, объясняющую эти результаты.
5. При скрещивании двух чистых линий гороха получены следующие результаты:
| Урожайность семян, ц/га | Высота растений, см | |
| Р1 | 25,0 | |
| Р2 | 17,6 | |
| F1 | 26,2 |
Вычислите гипотетический и истинный гетерозис по урожаю и высоте растений.
6. При скрещивании двух инбредных линий подсолнечника получены результаты:
| Масса 1000 зерен, г | Число зерен в корзинке, шт. | |
| Р1 | ||
| Р2 | ||
| F1 |
Проявился ли гетерозис в опытах по изучаемым признакам? Вычислите значение этих признаков в популяции F2.
7. Две инцухт-линии кукурузы, различающиеся по числу рядов зерен в початке и высоте прикрепления початка, были скрещены между собой. Выражение указанных признаков у родительских форм и гибридов F1 представлено ниже:
| Число рядов зерен в початке, шт. | Высота прикрепления початка, см | |
| Р1 | ||
| Р2 | ||
| F1 |
Определите гетерозис изучаемых признаков и возможное их значение в F2.
8. У перекрестноопыляющихся культур (например, у трав) раннецветущие растения скрещиваются с раннецветущими, а поздноцветущие – с поздноцветущими. При этом из популяции элиминируют очень раннецветущие генотипы из-за низкой завязываемости семян. Объясните, является ли данная ситуация типичным спонтанным ассортивным скрещиванием?
9. Предположим, что в культуру вводят две дикие формы, одна из которых принадлежит к самоопыляющимся, другая – к перекрестноопыляющимся культурам. Какую из этих форм можно быстрее разложить на линии? Объясните.
10. Отобранные с разными элементами продуктивности линии сои были скрещены между собой. Характеристика родительских форм и гибридов первого поколения представлена ниже.
| Число продуктивных узлов, шт. | Число зерен в бобе, шт. | |
| Р1 | 2,3 | |
| Р2 | 2,6 | |
| F1 | 2,5 |
Проявился ли гетерозис по изучаемым признакам? Какое число продуктивных узлов и число зерен в бобе ожидается в F2?
Поиск по сайту: