АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
Кровь как внутренняя среда организма
| Рис.2.2. Основные составные части крови. |
Кровь состоит из плазмы и клеток (форменных элементов) — эритроцитов, лейкоцитов и тромбоцитов, находящихся во взвешенном состоянии (рис.2.2.). Поскольку плазма и клеточные элементы имеют разобщенные источники регенерации, кровь часто выделяют в самостоятельный вид ткани.
Функции крови многообразны. Это, прежде всего, в обобщенном виде, функции транспорта или переноса газов и веществ, необходимых для жизнедеятельности клеток или подлежащих удалению из организма. К ним относятся: дыхательная, питательная, интегратив-но-регуляторная и экскреторная функции (см. главу 6).
Кровь выполняет в организме и защитную функцию, благодаря связыванию и нейтрализации токсических веществ, попадающих в организм, связыванию и разрушению инородных белковых молекул и чужеродных клеток, в том числе и инфекционного происхождения. Кровь является одной из основных сред, где осуществляются механизмы специфической защиты организма от чужеродных молекул и клеток, т.е. иммунитета.
Кровь участвует в регуляции всех видов обмена веществ и температурного гомеостазиса, является источником всех жидкостей, секретов и экскретов организма. Состав и свойства крови отражают сдвиги, происходящие в других жидкостях внутренней среды и клетках, в связи с чем исследования крови являются важнейшим методом диагностики.
Количество или объем крови у здорового человека находится в пределах 6-8 % массы тела (4 — 6 литров). Это состояние носит название нормоволемия. После избыточного приема воды объем крови может повышаться {гиперволемия), а при тяжелой физической работе в жарких цехах и избыточном потоотделении — падать (ги-поволемия).

|
| Рис.2.3. Определение ге-матокрита. |
Поскольку кровь состоит из клеток и плазмы, общий объем крови также складывается из объема плазмы и объема клеточных элементов. Часть объема крови, приходящаяся на клеточную часть крови, получила название гематокрит (рис. 2.3.). У здоровых мужчин гематокрит находится в пределах 44-48%, а у женщин — 41-45%. Благодаря наличию многочисленных механизмов регуляции объема крови и объема плазмы (волюморецепторные рефлексы, жажда, нервные и гуморальные механизмы изменения всасывания и выделения воды и солей, регуляция белкового состава крови, регуляция эритропоэза и др.) гематокрит является относительно жесткой гомеостатической константой и его длительное и стойкое изменение возможно лишь в условиях высокогорья, когда приспособление к низкому парциальному давлению кислорода усиливает эритропоэз и, соответственно, повышает долю объема крови, приходящуюся на клеточные
элементы. Нормальные величины гематокрита и, соответственно, объема клеточных элементов называют нормоцитемией. Увеличение объема, занимаемого клетками крови, называют полицитемией, а уменьшение — олигоцитемией.
Физико-химические свойства крови и плазмы. Функции крови во многом определяются ее физико-химическими свойствами, среди которых наибольшее значение имеют осмотическое давление, онко-тическое давление и коллоидная стабильность, суспензионная устойчивость, удельный вес и вязкость.
Осмотическое давление крови зависит от концентрации в плазме крови молекул растворенных в ней веществ (электролитов и неэлектролитов) и представляет собой сумму осмотических давлений содержащихся в ней ингредиентов. При этом свыше 60% осмотического давления создается хлористым натрием, а всего на долю неорганических электролитов приходится до 96% от общего осмотического давления. Осмотическое давление является одной из жестких гомеостатических констант и составляет у здорового человека в среднем 7,6 атм с возможным диапазоном колебаний 7,3-8,0 атм. Если жидкость внутренней среды или искусственно приготовленный раствор имеет такое же осмотическое давление, как нормальная плазма крови, подобную жидкую среду или раствор называют изотоническим. Соответственно, жидкость с более высоким осмотическим давлением называется гипертонической, а с более низким — гипотонической.
Осмотическое давление обеспечивает переход растворителя через полунепроницаемую мембрану от раствора менее концентрированного к раствору более концентрированному, поэтому оно играет важную роль в распределении воды между внутренней средой и клетками организма. Так, если тканевая жидкость будет гипертонической, то вода будет поступать в нее с двух сторон — из крови и из клеток, напротив, при гипотоничности внеклеточной среды вода переходит в клетки и кровь.
Аналогичную реакцию можно наблюдать со стороны эритроцитов крови при изменении осмотического давления плазмы: при гипер-тоничности плазмы эритроциты, отдавая воду, сморщиваются, а при гипотоничности плазмы набухают и даже лопаются. Последнее, используется в практике для определения осмотической стойкости эритроцитов. Так, изотоничным плазме крови является 0,89% раствор NaCl. Помещенные в этот раствор эритроциты не изменяют формы. В резко гипотоничных растворах и, особенно, воде эритроциты набухают и лопаются. Разрушение эритроцитов носит название гемолиз, а в гипотоничных растворах — осмотический гемолиз. Если приготовить ряд растворов NaCl с постепенно уменьшающейся концентрацией поваренной соли, т.е. гипотоничные растворы, и помешать в них взвесь эритроцитов, то можно найти ту концентрацию гипотоничного раствора, при котором начинается гемолиз и единичные эритроциты разрушаются или гемолизируются. Эта концентрация NaCl характеризует минимальную осмотическую резистент-
61
ность эритроцитов (минимальный гемолиз), которая у здорового человека находится в пределах 0,5-0,4 (% раствора NaCl). В более гипотонических растворах все более количество эритроцитов гемо-лизируется и та концентрация NaCl, при которой все эритроциты будут лизированы, носит название максимальной осмотической ре-зистентности (максимальный гемолиз). У здорового человека она колеблется от 0,34 до 0,30 (% раствора NaCl).
Механизмы регуляции осмотического гомеостазиса изложены в главе 12.
Онкотическое давление и коллоидная стабильность белков плазмы крови. Онкотическим давлением называют осмотическое давление, создаваемое белками в коллоидном растворе, поэтому его еще называют коллоидно-осмотическим. Ввиду того, что белки плазмы крови плохо проходят через стенки капилляров в тканевую микросреду, создаваемое ими онкотическое давление обеспечивает удержание воды в крови. Если осмотическое давление, обусловленное солями и мелкими органическим молекулами, из-за проницаемости гистогематичес-ких барьеров одинаково в плазме и тканевой жидкости, то онкотическое давление в крови существенно выше. Кроме плохой проницаемости барьеров для белков, меньшая их концентрация в тканевой жидкости связана с вымыванием белков из внеклеточной среды током лимфы. Таким образом, между кровью и тканевой жидкостью существует градиент концентрации белка и, соответственно, градиент он-котического давления. Так, если онкотическое давление плазмы крови составляет в среднем 25-30 мм рт.ст., а в тканевой жидкости — 4-5 мм рт.ст., то градиент давления равен 20-25 мм рт.ст. Поскольку из белков в плазме крови больше всего содержится альбуминов, а молекула альбумина меньше других белков и его моляльная концентрация поэтому почти в 6 раз выше, то онкотическое давление плазмы создается преимущественно альбуминами. Снижение их содержания в плазме крови ведет к потере воды плазмой и отеку тканей, а увеличение — к задержке воды в крови.
Коллоидная стабильность плазмы крови обусловлена характером гидратации белковых молекул и наличием на их поверхности двойного электрического слоя ионов, создающего поверхностный или фи-потенциал. Частью фи-потенциала является электрокинетический (дзета) потенциал. Дзета-потенциал — это потенциал на границе между коллоидной частицей, способной к движению в электрическом поле, и окружающей жидкостью, т.е. потенциал поверхности скольжения частицы в коллоидном растворе. Наличие дзета-потенциала на границах скольжения всех дисперсных частиц формирует на них одноименные заряды и электростатические силы отталкивания, что обеспечивает устойчивость коллоидного раствора и препятствует агрегации. Чем выше абсолютное значение этого потенциала, тем больше силы отталкивания белковых частиц друг от друга. Таким образом, дзета-потенциал является мерой устойчивости коллоидного раствора. Величина этого потенциала существенно выше у альбуминов плазмы, чем у других белков. Поскольку альбуминов
62
в плазме значительно больше, коллоидная стабильность плазмы крови преимущественно определяется этими белками, обеспечивающими коллоидную устойчивость не только других белков, но и углеводов и липидов.
С коллоидной стабильностью белков плазмы связаны и суспензионные свойства крови, т.е. поддержание клеточных элементов во взвешенном состоянии. Величина суспензионных свойств крови может быть оценена по скорости оседания эритроцитов (СОЭ) в неподвижном объеме крови.
Таким образом, чем выше содержание альбуминов по сравнению с другими, менее стабильными коллоидными частицами, тем больше и суспензионная способность крови, поскольку альбумины адсорбируются на поверхности эритроцитов. Наоборот, при повышении в крови уровня глобулинов, фибриногена, других крупномолекулярных и нестабильных в коллоидном растворе белков, скорость оседания эритроцитов нарастает, т.е. суспензионные свойства крови падают. В норме СОЭ у мужчин 4-10 мм/ч, а у женщин — 5-12 мм/ч.
Вязкость — это способность оказывать сопротивление течению жидкости при перемещениях одних частиц относительно других за счет внутреннего трения. В связи с этим, вязкость крови представляет собой сложный эффект взаимоотношений между водой и макромолекулами коллоидов с одной стороны, плазмой и форменными элементами — с другой. Поэтому вязкость плазмы и вязкость, цельной крови существенно отличаются: вязкость плазмы в 1,8 — 2,5 раза выше, чем воды, а вязкость крови выше вязкости воды в 4- 5 раз. Чем больше в плазме крови содержится крупномолекулярных белков, особенно фибриногена, липопротеинов, тем выше вязкость плазмы. При увеличении количества эритроцитов, особенно их соотношения с плазмой, т.е. гематокрита, вязкость крови резко возрастает. Повышению вязкости способствует и снижение суспензионных свойств крови, когда эритроциты начинают образовывать агрегаты. При этом отмечается положительная обратная связь — повышение вязкости, в свою очередь, усиливает агрегацию эритроцитов — что может вести к порочному кругу. Поскольку кровь — неоднородная среда и относится к неньютоновским жидкостям, для которых свойственна структурная вязкость, постольку снижение давления потока, например, артериального давления, повышает вязкость крови, а при повышении давления из-за разрушения структурированности системы — вязкость падает.
Еше одной особенностью крови как системы, обладающей наряду с ньютоновской и структурной вязкостью, является, эффект Фареу-са-Линдквиста. В однородной ньютоновской жидкости, согласно закону Пуазейля, с уменьшением диаметра трубки повышается вязкость. Кровь, которая является неоднородной неньютоновской жидкостью, ведет себя иначе. С уменьшением радиуса капилляров менее 150 мк вязкость крови начинает снижаться. Эффект Фареуса-Линд-квиста облегчает движение крови в капиллярах кровеносного русла. Механизм этого эффекта связан с образованием пристеночного слоя плазмы, вязкость которой ниже, чем у цельной крови, и миграцией
эритроцитов в осевой ток. С уменьшением диаметра сосудов толщина пристеночного слоя не меняется. Эритроцитов в движущейся по узким сосудам крови становится по отношению к слою плазмы меньше, т.к. часть из них задерживается при вхождении крови в узкие сосуды, а находящиеся в своем токе эритроциты двигаются быстрее и время пребывания их в узком сосуде уменьшается.
Вязкость крови прямо пропорционально сказывается на величине общего периферического сосудистого сопротивления кровотоку, т.е. влияет на функциональное состояние сердечно-сосудистой системы.
Удельный вес крови у здорового человека среднего возраста составляет от 1,052 до 1,064 и зависит от количества эритроцитов, содержания в них гемоглобина, состава плазмы. У мужчин удельный вес выше, чем у женщин за счет разного содержания эритроцитов. Удельный вес эритроцитов (1,094-1,107) существенно выше, чем у плазмы (1,024-1,030), поэтому во всех случаях повышения гемато-крита, например, при сгущении крови из-за потери жидкости при потоотделении в условиях тяжелой физической работы и высокой температуры среды, отмечается увеличение удельного веса крови.
Состав плазмы крови. Плазма крови состоит из воды (около 90% массы), низко молекулярных соединений органического и неорганического происхождения — солей или электролитов, углеводов, ли-пидов, органических кислот и оснований, промежуточных продуктов обмена как содержащих азот, так и неазотистого происхождения, витаминов (около 2% массы) и белков, на долю которых приходится до 8% массы плазмы.
Электролитный состав плазмы важен для поддержания ее осмотического давления, кислотно-щелочного состояния, функций клеточных элементов крови и сосудистой стенки, активности ферментов, процессов свертывания крови и фибринолиза. Поскольку плазма крови постоянно обменивается электролитами с микросредой клеток, содержание в ней электролитов в значительной мере определяет и фундаментальные свойства клеточных элементов органов — возбудимость и сократимость, секреторную активность и проницаемость мембран, биоэнергетические процессы. Содержание основных электролитов в плазме крови, эритроцитах и тканевой микросреде представлено в табл.2.1. Из таблицы видно, что содержание натрия и калия в плазме и эритроцитах отличается также, как и в других клетках и внеклеточной среде (глава 1), и, соответственно, обусловлено различиями проницаемости мембран и работой К- Na- насосов клеток. Часть катионов плазмы связана с анионами органических кислот и белков, что играет роль в поддержании кислотно-щелочного состояния и необходимо для реализации функций белков.
Отличается в плазме и эритроцитах содержание и ряда анионов, прежде всего хлора и бикарбоната. Эти различия обусловлены обменом этих анионов между эритроцитами и плазмой в капиллярах легких и тканей при дыхании.
Содержание натрия и калия в плазме крови — жесткие гомеоста-тические константы, зависящие от баланса процессов поступления и
64
Таблица 2.1. Содержание электролитов в плазме крови, эритроцитах и микросреде тканей (ммоль/л) у человека
| Ионы | Плазма | Эритроциты | Микросреда клеток |
| Катионы | |||
| Na* | 135-150 | 10-25 | 142-144 |
| К+ | 4,0-5,5 | 95-110 | 4,0-5,0 |
| Са+ | 2,2-2,5 | 0,12-0,30 | 0,8-1,2 |
| Мg* | 0,5-0,9 | 1,7-2,3 | 0,75-1,2 |
| Анионы | |||
| CI | 90-110 | 40-60 | 100-120 |
| НСО3 | 23-33 | 13-18 | 28-32 |
| HPOf | 0,8-1,2 | 38-48 | 0,8-1,2 |
| so42- | 0,4-0,6 | 6-8 | 0,4-0,6 ■ |
выведения ионов, а также их перераспределения между клетками и внеклеточной средой. Регуляция гомеостазиса этих катионов осуществляется изменениями поведения (большее или меньшее потребление соли) и системами гуморальной регуляции (см.главу 3), среди которых основное значение имеют ренин- ангиотензин- альдостеро-новая система и натриуретический гормон предсердий (см.главу 5). Жесткой гомеостатической константой является и концентрация кальция в плазме крови. Кальций содержится в двух формах: связанной (с белками, в комплексных соединениях, малорастворимых солях) и свободной, ионизированной (Са++). Основные биологические эффекты кальция обусловлены его ионизированной формой. В цитозоле клеток ионизированного кальция содержится мало, но его количество чрезвычайно тонко регулируется, поскольку этот катион является важнейшим регулятором обменных процессов и функций клеток. Поступление кальция в клетки из внеклеточной среды связано с его уровнем в микросреде и плазме крови, хотя в большей степени зависит от специальных транспортных мембранных механизмов (каналов, насосов, переносчиков). В клеточном цитозоле ионизированный кальций связывается с белками, а также удаляется с помощью специальных Са-насосов во внутриклеточные депо (митохондрии, цитоплазматический ретикулум) и наружу в микросреду клеток. Содержащийся в плазме крови ионизированный кальций помимо того, что является источником для транспорта внутрь клеток, необходим для обеспечения физико-химических свойств плазменных белков, активности ферментов, например, для реализации механизмов свертывания крови. Регуляция уровня ионизированного кальция в плазме крови осуществляется специальной гуморальной
65
системой, включающей ряд кальций-регулирующих гормонов: око
лощитовидных желез (паратирин), щитовидной железы (кальцитонин
и его аналоги), почек (кальцитриол).,
В плазме крови содержится и большое число различных микроэлементов, называемых так из- за очень малых концентраций. Как минимум 15 микроэлементов, содержащихся в плазме крови, например, медь, кобальт, марганец, цинк, хром, стронций и др., играют важную роль в процессах метаболизма клеток и обеспечении их функций, поскольку входят в состав ферментов, катализируют их действие, участвуют в процессах образования клеток крови и гемоглобина (гемопоэзе) и др.
Из веществ органической природы в плазме крови находятся азо-тосодержащие продукты белкового катаболизма (мочевина, аминокислоты, мочевая кислота, креатин, креатинин, индикан), получившие название остаточного или небелкового азота. Величина остаточного азота (в норме 14,3-28,6 ммоль/л) отражает не столько интенсивность катаболизма белка, сколько эффективность выделения продуктов белкового обмена через почки. При нарушениях экскреторной функции почек повышение остаточного азота крови является важным диагностическим показателем.
Необходимым для жизнедеятельности организма является содержание в плазме крови углеводов, из которых более 90% приходится на глюкозу. Благодаря высокой растворимости в воде, хорошей способности к мембранному транспорту и легкости использования в метаболических путях, глюкоза, для многих клеток организма является главным источником энергии. Содержание глюкозы в артериальной крови выше, чем в венозной, так как она непрерывно используется клетками тканей. У здорового человека в венозной крови содержится 3,6-6,9 ммоль/л глюкозы, причем колебания ее уровня преимущественно связаны с временем приема пищи и всасыванием из желудочно-кишечного тракта. В целом уровень глюкозы в крови зависит от соотношения следующих факторов: всасывания из желудочно-кишечного тракта, поступления из депо (гликоген печени), новообразования из аминокислот и жирных кислот (глюконеогенез), утилизации тканями и депонирования в виде гликогена. Гомеостазис глюкозы отражает особенности углеводного обмена в организме и регулируется вегетативной нервной системой и многочисленными сахар-регулирующими гормонами (инсулин, глюкагон, адреналин, глюкокортикоиды и др.).
Важную роль в реализации питательной функции крови играют содержащиеся в плазме липиды и белки.
Белки плазмы крови. Общее число белков плазмы крови составляет около 200, из них 70 выделены в чистом виде. Общее содержание белка в крови колеблется в норме от 65 до 85 г/л. Основными плазменными белками являются альбумины (38-50 г/л), глобулины (20-30 г/л) и фибриноген (2-4 г/л). Таким образом, больше всего в плазме крови содержится альбуминов, и для оценки белкового состава плазмы в клинике обычно определяют альбумино/гло-
66
булиновый показатель или белковый коэффициент крови, составляющий у здоровых взрослых людей 1,3-2,2. С помощью электрофореза, т.е. передвижения белковых частиц в электрическом поле, удается выделить так называемые белковые фракции, каждая из которых, кроме альбуминов, образована большим количеством разных по составу белковых молекул. Содержание в плазме основных белковых фракций приведено в табл.2.2. Выявление белковых фракций основано лишь на физико-химических свойствах белков, а не на физиологическом их значении, поэтому в одну и ту же фракцию попадают белки с разными функциональными свойствами. Наиболее же точную информацию о белковом составе плазмы можно получить определяя содержание индивидуальных белков.
Таблица 2.2. Основные белковые фракции плазмы крови человека
| Фракции | Концентрации (г/л) |
| Общий белок | 65-85 |
| Альбумины | 38-50 |
| Альфа-1-Глобулины | 1,4-3,0 |
| Альфа-2-Глобулины | 5,6-9,1 |
| Бета-Глобулины | 5,4-9,1 |
| Гамма-Глобулины | 9,1-14,7 |
| Фибриноген | 2,0-4,0 |
Альбумины — самая однородная фракция белков плазмы. Основная их функция заключается в поддержании онкотического давления. Кроме того альбумины служат резервом аминокислот для белкового синтеза и выполняют тем самым питательную функцию. Благодаря большой поверхности мицелл и их высокому отрицательному заряду, альбумины обеспечивают стабильность коллоидного раствора и суспензионные свойства крови, адсорбируют на своей поверхности и транспортируют вещества не только эндогенного, но и экзогенного происхождения. Так, альбумины переносят неэстери-фицированные жирные кислоты, билирубин, стероидные гормоны, соли желчных кислот, а также, пенициллин, сульфаниламиды, ртуть. Альбумины частично связывают гормон щитовидной железы тироксин и значительную часть ионов кальция.
Альфа-глобулины включают гликопротеины, т.е. белки, связанные с углеводами (2/3 всей глюкозы плазмы циркулирует в составе гликопротеинов), а также ингибиторы протеолитических ферментов, транспортные белки для гормонов, витаминов и микроэлементов. К альфа-глобулинам относятся: эритропоэтин — гуморальный стимулятор кроветворения; плазминоген — предшественник фермента, растворяющего свернувшуюся кровь; протромбин — один из факторов свертывания крови и т.д. Альфа-глобулины осущест-
67
вляют транспорт липидов, участвуя в образовании липопротеидных комплексов, в составе которых переносятся триглицериды, фосфо-липиды, холестерин и сфингомиелины.
Бета-глобулины — самая богатая липидами фракция белка. Находясь в составе липопротеидов, эти белки содержат 3/4 всех липидов плазмы крови, в том числе фосфолипиды, холестерин и сфингомиелины. К этой белковой фракции относятся белок транс-феррин, обеспечивающий транспорт железа, большая часть белков системы комплемента, многие факторы свертывания крови.
Гамма- глобулины называют также иммуноглобулинами, поскольку в эту фракцию входят антитела или иммуноглобулины (Ig) 5 классов: IgA, IgG, IgM, IgD, IgE.
В общем функции белков плазмы крови сводятся к обеспечению: 1) коллоидно-осмотического и водного гомеостаза, 2) агрегатного состояния крови и ее реологических свойств (вязкость, свертываемость, суспензионные свойства), 3) кислотно-щелочного гомеостаза, 4) иммунного гомеостаза, 5) транспортной функции крови и 6) питательной функции крови, как резерв аминокислот.
2.2. Интерстициальное пространство и микросреда клеток.
Интерстициальное пространство представляет собой сеть колла-геновых и эластических волокон, ячейки которой заполнены геле-образным веществом, состоящим из белков, полисахаридов типа гли-козаминогликанов-гиалуроновой кислоты, хондроитинсульфатов А, В и С, минеральных солей и воды. Коллаген — это белковые волокна, образуемые фиброцитами соединительной ткани. Масса коллагено-вых волокон очень велика и составляет порядка 6% массы тела, а обшая поверхность волокон превышает миллион квадратных метров. Такая структура сети создает своеобразную коллагеновую "губку", способную накапливать воду и электролиты, особенно натрий. Связывание воды и электролитов коллагеном увеличивается при появлении в интерстициальном пространстве избытка Н-ионов, например, молочной кислоты. Полисахариды типа гликозаминогликанов синтезируются фибробластами, что обеспечивает постоянство отрицательных зарядов (анионов) в интерстиции. Активирует синтез гликозаминогликанов инсулин, подавляют — катехоламины, а тиреоид-ные гормоны способствуют катаболизму этих полисахаридов.
Белки и полисахаридные анионы образуют в интерстиции комплексы, называемые муко- и гликопротеины. Указанные компоненты формируют коллоидную или гелеподобную фазу интерстиция, которая способна связывать воду и набухать, благодаря высокой гидрофильности, или освобождаться от воды под влиянием местно действующих ферментов и биологически активных веществ (гиалу-ронидаза, гепарин, гистамин и др.), как бы передавая воду следующим ячейкам основного вещества. Таким образом, эта фаза межуточного вещества осуществляет избирательно замедленный транспорт микромолекул, поступивших в интерстициальное пространство.
68
Эта фаза способна также замедлять продвижение молекул с увеличением их массы, что ограничивает транспорт крупномолекулярных белков, поступивших в интерстииий из крови или клеток. Вторая фаза — водная, в виде тонких "каналов" вдоль фибриллярных волокон, обеспечивает свободный транспорт микромолекул. В обеих фазах интерстициального пространства содержится много воды. В общем, у человека массой 70 кг в интерстициальном пространстве содержится в среднем 10,6 л жидкости. Электролитный состав тканевой жидкости приведен в табл.2.1.
Вода интерстициального пространства постоянно обменивается с плазмой крови кровеносных капилляров. В артериальной части капилляра гидростатическое давление крови превышает онкотическое давление белков плазмы, гидростатическое давление тканевой жидкости и вода фильтруется через гистогематический барьер в интерс-тициальное пространство. К венозному концу капилляра гидростатическое давление крови снижается, из-за выхода воды в ткань несколько повышается концентрация белков в плазме и онкотическое давление становится выше гидростатического, что обеспечивает обратное поступление воды из тканей в кровь (рис.2.4.).
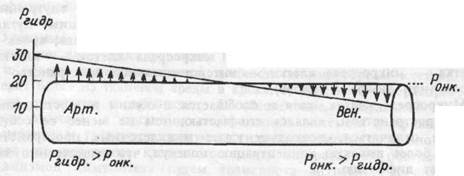
Рис. 2.4. Роль гидростатического давления крови в транскапиллярном обмене воды.
Арт. и Вен. — артериальный и венозный участки капилляра. Стрелки показывают направление и интенсивность движения воды.
Меньшие концентрации белка в интерстициальном пространстве обуславливают и меньшие величины онкотического давления. Давление тканевой жидкости широко варьирует от -6 до +12 мм рт.ст. Величина этого давления, в основном, характеризует водную фазу интерс-тициального пространства и зависит от ряда причин — скорости фильтрации воды из кровеносных капилляров, количества воды, связываемой коллоидной фазой, скорости оттока воды в лимфатические капилляры. В свою очередь давление в интерстициальном пространстве определяет интенсивность фильтрации воды и лимфооттока.
Интерстициальное пространство содержит ряд клеток соединительной ткани — фибробласты и фиброциты, мастоциты или тучные клетки, макрофаги и лимфоциты. Эти клетки регулируют состояние интерстициального пространства, микросреды клеток, кровеносных и лимфатических капилляров, благодаря образованию биологически-активных веществ — ферментов, гепарина, биогенных аминов. Собственно процессы метаболизма интерстициального пространства протекают в микроячейке: фиброциты, фибробласты, мастоциты — основное вещество — коллагеновые волокна (коллоидная фаза) — вода и электролиты (водная фаза). Клетки интерстициального пространства осуществляют фагоцитоз, участвуют в механизмах иммунитета.
Микросредой клеток называют часть внеклеточной внутренней среды или интерстициального пространства, непосредственно прилегающую к поверхности клеток. Это своего рода "атмосфера" клетки, имеющая очень малую толщину (порядка 20 нм), но играющая основную роль в обмене веществ через мембрану клетки. Состав и свойства микросреды клеток во многом одинаковы с интерстици-альным пространством, поскольку обмен между ними происходит путем простой диффузии. Вместе с тем, если интерстициальное пространство в большей мере связано с транспортом веществ из крови, то микросреда клеток в большей мере связана с метаболическими процессами в клетках. Транспорт веществ во внутренней среде организма может осуществляться по следующим путям: кровь — интерстициальное пространство — лимфа — кровь; кровь — интерстициальное пространство — микросреда клеток — клетки; клетка — микросреда клетки — интерстициальное пространство — кровь (или лимфа — кровь).
Микросреда клеток, хотя и сообщается с общим интерстициаль-ным пространством, являясь его частью, тем не менее ее состав должен отличаться, поскольку узкие межклеточные пространства имеют более высокую концентрацию молекул, что существенно затрудняет диффузию.
Обмен между микросредой клеток и общим интерстициальным пространством происходит, в основном, за счет "конвективного" транспорта (т.е. перемещения воды и молекул под влиянием градиентов сил гидростатического, онкотического и осмотического давлений, электростатических и электрокинетических потенциалов). Клеточная микросреда богата продуктами метаболизма клеток и протео-лиза отмерших структур. Полисахариды в этой околоклеточной микросреде образуют гликокалике, располагающийся на поверхности клеточной мембраны и существенно замедляющий процессы диффузии в клетку и из клетки. Гликокаликс участвует в трансмембранном обмене ионов, явлениях адгезии (прилипания) между клетками, процессах тканевой дифференцировки, функционирования базальных мембран, иммунологических реакциях.
В микросреде клеток могут накапливаться аминокислоты и жирные кислоты, формируя тем самым резервный фонд, необходимый для пластических и энергетических процессов в клетке. Микросреда служит пространством переноса медиаторов и гормонов, тем самым
70
активно участвуя в процессах регуляции клеточных функций и метаболизма. Изменение свойств микросреды клеток меняет концентрацию или время нахождения этих гуморальных регуляторов у рецепторов клеточной мембраны, что ведет к изменению выраженности и длительности регуляторного влияния, а следовательно, и эффективности регуляции, как таковой.
Мембрана клеток способна фиксировать не только ферменты микросреды, но и циркулирующие во внутренней среде антигены. Поэтому микросреда клеток является важнейшим участком развертывания процессов иммунитета.
Поиск по сайту: