АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
Законы раздражения. Параметры возбудимости
Понятие о раздражимости, возбудимости и возбуждении. Классификация раздражителей.
Раздражимость - это способность клеток, тканей, организма в целом переходить под воздействием факторов внешней или внутренней среды из состояния физиологического покоя в состояние активности. Состояние активности проявляется изменением: физиологических параметров клетки, ткани организма, например изменением метаболизма.
Возбудимость - это способность живой ткани отвечать на раздражение активной специфической реакцией — возбуждением, т.е. генерацией нервного импульса, сокращением, секрецией. Т.е. возбудимость характеризует специализированные ткани - нервную, мышечные, железистые, которые называются возбудимыми. Возбуждение - это комплекс процессов реагирования возбудимой ткани на действие раздражителя, проявляющийся изменением мембранного потенциала, метаболизма и т.д.. Возбудимые ткани обладают проводимостью-это способность ткани проводить возбуждение. Наибольшей проводимостью обладают нервы и скелетные мышцы. Раздражитель - это фактор внешней или внутренней среды действующий на живую ткань. Процесс воздействия раздражителя на клетку, ткань, организм называется раздражением. Все раздражители делятся на следующие группы:
1. По природе
а) физические (электричество, свет, звук,механические воздействия и т.д.)
б) химические (кислоты, щелочи, гормоны и т.д.)
в) физико-химические (осмотическое давление, парциальное давление газов и т.д.)
г) биологические (пища для животного, особь другого пола)
д) социальные (слово для человека).
2. По месту воздействия
а) внешние (экзогенные)
б) внутренние (эндогенные)
3. По силе:
а) подпороговые (не вызывающие ответной реакции)
б) пороговые (раздражители минимальной, силы, при которой возникает возбуждение)
в) сверхпороговые (силой выше пороговой)
4. По физиологическому характеру:
а) адекватные (физиологичные для данной клетки или рецептора, которые, приспособились к нему в процесс эволюции, например, свет для фоторецепторов глаза).
б) неадекватные
Если реакция на раздражитель является рефлекторной, то выделяют также
а) безусловно-рефлекторные раздражители
б) условно-рефлекторные
Законы раздражения. Параметры возбудимости.
Реакция клеток, тканей на раздражитель определяется законами раздражения
1. Закон "все или ничего": При допороговых раздражениях клетки в ткани ответной реакции не возникает. При пороговой силе раздражителя развивается максимальная ответная реакция, поэтому увеличение силы раздражения выше пороговой не сопровождается ее усилением. В соответствии с этим законом реагирует на раздражения одиночное нервное и мышечное волокно, сердечная мышца.
2. Закон силы: Чем больше сила раздражителя, тем сильнее ответная реакция. Однако выраженность ответной реакции растет лишь до определенного максимума. Закону силы подчиняется целостная скелетная, гладкая мышца, ТАК как они состоят из многочисленных мышечных клеток, имеющих различную возбудимость.
3. Закон силы-длительности. Между силой и длительностью действия раздражителя имеется определенная взаимосвязь. Чем сильнее раздражитель, тем меньшее время требуется для возникновения ответной реакции. Зависимость между пороговой силой и необходимой длительностью раздражения отражается кривой силы длительности. По этой кривой можно определить ряд параметров возбудимости.
а) Порог раздражения - это минимальная сила раздражителя, при которой возникает возбуждение.
б) Реобаза - это минимальная сила раздражителя, вызывающая возбуждение при его действии в течение неограниченно долгого времени. На практике порог и реобаза имеют одинаковый смысл. Чем ниже порог раздражения или меньше реобаза, тем выше возбудимость ткани.
в) Полезное время - минимальное время действия раздражителя силой в одну реобазу за которое возникает возбуждение.
г) Хронаксия - это минимальное время действия раздражителя силой в две реобазы, необходимое для возникновения возбуждения.
Этот параметр предложил рассчитывать Л. Лапик для более точного определения показателя времени на кривой силы-длительности. Чем короче полезное время или хронаксия тем выше возбудимость и наоборот. В клинической практике реобазу и хронаксию определяют с помощью метода хронаксимстрии для исследования возбудимости нервных стволов.
4. Закон градиента или аккомодации. Реакция ткани на раздражение зависит от его градиента, т.е. чем быстрее нарастает сила раздражителя во времени тем быстрее возникает ответная реакция. При низкой скорости нарастания силы раздражителя растет порог раздражения. Поэтому если сила раздражителя, возрастает очень медленно возбуждения не будет. Это явление называется аккомодацией. Физиологическая лабильность (подвижность) - это большая или меньшая частота реакций, которыми может отвечать ткань на ритмическое раздражение. Чем быстрее восстанавливается ее возбудимость после очередного раздражения, тем Выше ее лабильность. Определение лабильности предложено Н.Е.Введенским. Наибольшая, лабильность у нервов, наименьшая у сердечной мышцы.
Действие постоянного тока на возбудимые ткани.
В первые закономерности действия постоянного тока на нерв нервно-мышечного препарата исследовал в 19веке Пфлюгер. Он установил, что при замыкании цепи постоянного тока, под отрицательным.. электродом т е. катодом
•возбудимость повышается, а под положительным - анодом снижается. Это называется законом действия постоянного Тока. Изменение возбудимости ткани (например: нерва) под действием постоянного тока в области анода или катода называется физиологическим электротоном. В настоящее время установлено, что под действием отрицательного электрода - катода потенциал мембраны клеток снижается. Это явление называется физическим катэлектротоном, Под положительным - анодом, он возрастает. Возникает физический катэлектртон. Так как, под катодом мембранный потенциал приближается к критическому уровню деполяризации, возбудимость клеток и тканей повышается. Под анодом мембранный потенциал возрастает и удаляется от критического уровня деполяризации, поэтому возбудимость клетки, ткани падает. Следует отметить, что при очень кратковременном действии постоянного тока (1 мсек и менее)МП не успевает измениться, поэтому не изменяется и возбудимость ткани под электродами.
Постоянный ток широко используется в клинике для лечения и диагностики. Например, с помощью него производится электростимуляция нервов и мышц, физипроцедуры: ионофорез и гальванизация. Строение и функции цитоплазматической мембраны клеток. Цитоплазматическая клеточная мембрана состоит из трех слоев: наружного белкового, среднего бимолекулярного слоя липидов и внутреннего белкового. Толщина мембраны 7.5'-10 нм. Бимолекулярный слой липидов является матриксом мембраны. Липидные молекулы его обоих слоев взаимодействуют с белковыми молекулами. погруженными в них. От 60 до 75% липидов мембраны составляют фосфолипиды. 15- 30% холестерина. Белки представлены в основном гликопротеинами. Различают интегральные белки, пронизывающие всю мембрана и периферические, находящиеся на наружной или внутренней поверхности. Интегральные белки образуют ионные каналы, обеспечивающие обмен определенных ионов между вне- и внутриклеточной жидкостью. Они также являются ферментами, осуществляющими противоградиентный перенос ионов через мембрану. Периферическими белками являются хеморецепторы наружной поверхности мембраны, которые могут взаимодействовать
ФУНКЦИИ мембраны:
1. Обеспечивает целостность клетки, как структурной единицы ткани.
2. Осуществляет обмен ионов между цитоплазмой и внеклеточной жидкостью,
3. Обеспечивает активный транспорт ионов и других веществ в клетку и из нее
4. Производит восприятие и переработку информации поступающей к клетке в виде химических и электрических
Механизмы возбудимости клеток. Ионные каналы мембраны. Механизмы возникновения мембранного потенциала (М П) и потенциалов действия /П.Л)
(в основном, передаваемая в организме информация имеет вид электрических сигналов (например нервные импульсы). Впервые наличие животного электричества установил физиолог Л Гальвани в 1736 г.. С целью исследования атмосферного электричества он подвешивал нервно-мышечные препараты лапок лягушек на медном крючке. Когда эти лапки касались железных перил балкона, происходило сокращение. мышц. Это свидетельствовало о действии какого-то электричества на нерв нервно-мышечного препарата. Гальвани посчитал, что это обусловлено наличием электричества в самих живых тканях. Однако А. Вольта установил, что источником электричества является место контакта двух разнородных металлов -.меди и железа. В физиологии первым классическим опытом Гальвани считается прикосновение к нерву нервно-мышечного препарата биметаллическим пинцетом, сделанным из меди и железа. Чтобы доказать свою правоту, Гальвани произвел второй опыт. Он набрасывал конец нерва, нннервируюшего нервно-мышеччый препарат, на разрез его мышцы. В результате возникало ее сокращение. Однако и этот опыт не убедил современников Гальвани. Поэтому другой итальянец Маттеучи произвел следующий эксперимент. Он накладывал нерв одного нервно-мышечного препарат лягушки на мышцу второго, которая сокращалась под действием раздражающего тока. В результате первый препарат тоже начинал сокращаться. Это свидетельствовало о передаче электричества (ПД) от одной мышце к другой. Наличие разности потенциалов между поврежденным и неповрежденным участками мышцы впервые точно установил, а 19 веке с помощью струнного гальванометра (амперметра) Маттеучи. Причем разрез имел отрицательный заряд, а поверхность мышцы положительный.
Классификация и структура ионных каналов цитоплазматической мембраны. Механизмы возникновения
мембранного потенциала и потенциалов действия.
.'Первый шаг в изучении причин возбудимости клеток сделал в своей работе "Теория мембранного равновесия" в 1924 г. английский физиолог Донанн. Он теоретически установил, что разность потенциалов внутри клетки и вне ее, т.г. потенциала покоя или МП, близка к калиевому равновесному потенциалу, Это потенциал, образующемуся на полупроницаемой мембране разделяющий растворы с разной концентрацией ионов калия, один из которых содержит крупные непроникающие анионы. Его расчеты уточнил Нернст. Он вывел уравнение диффузионного потенциала для калия он будет равен: Экспериментально механизмы возникновения разности потенциалов между внеклеточной жидкостью и цитоплазмой, а также возбуждения клеток установили в 1939 году в Кембридже Ходжкин и Хаксли. Они исследовали нервное гигантское волокно (аксон) кальмара и обнаружили, что внутриклеточная жидкость нейрона содержит 400 мМ калия. 50 мМ натрия, 100 мМ хлора очень мало кальция.
Во внеклеточной жидкости содержалось всего 10 мМ калия, 440 мМ, "натрия, 560 мМ хлора и 10-мМ кальция. Таким образом, внутри клеток имеется избыток калия, а вне их натрия и кальция. Это обусловлено тем,что в клеточною мембрану встроены ионные каналы, регулирующие проницаемость мембраны для ионов натрия, калия, кальция и хлора. Все ионные каналы подразделяются на следующие группы:
1. По избирательности:
а) Селективные, т.е. специфические. Эти каналы проницаемы для строго определенных ионов. б)Малоселективные, неспецифические, не имеющие определенной ионной избирательности: Их в мембране. небольшое количество.
2. По характеру пропускаемых ионов:
а) калиевые и натриевые
в) кальциевые
г)хлорные
3. По скорости инактивации, т.е. закрывания:
а) быстроинактивируюшиеся, т.е. быстро переходящие в закрытое состояние. Они обеспечивают быстро нарастающее снижение МП и такое же быстрое восстановление.
б) медленноинактирующиеся. Их открывание вызывает медленное снижение МП и медленное его восстановление.
4. По механизмам открывания:
а) потенциалзависямые, т.е. те, которые открываются при определенном уровне потенциала мембраны.
б) хемозависимые, открывающиеся при воздействии на хеморецепторы мембраны клетки физиологически
активных веществ (нейромедиаторов. гормонов и т. д).
В настоящее время установлено, что ионные каналы имеют следующее строение:
1.Селективный фильтр, расположенный в устье канала. Он обеспечивает прохождение через канал строго
определенных ионов.
2.Активационные ворота, которые открываются при определенном уровне мембранного потенциала или действии соответствующего ФАВ. Активационные ворота потенциалзависямых каналов имеется сенсор, который открывает их на определенном уровне МП.
3.Инактивационные ворота, обеспечивающие закрывания канала и прекращение проведения ионов по каналу на определенном уровне МП. (Рис). Неспецифические ионные каналы не имеют ворот.
Селективные ионные каналы могут находиться в трех состояниях, которые определяются положением активационных (м) и инактивационных (н) ворот (рис):
1. Закрытом, когда активационные закрыты, а инактивацинные открыты.
2. Активированном, и те и другие ворота открыты.
3. Инактивированном активационные ворота открыты, а инактивационные закрыты
Суммарная проводимость для того или иного иона определяется числом одновременно открытых соответствующих каналов. В состоянии покоя открыты только калиевые каналы, обеспечивающие поддержание определенного
мембранного потенциала и закрыты натриевые. Поэтому мембрана избирательно проницаема для калия и очень мало для ионов натрия и кальция, за счет имеющихся неспецифических каналов. Соотношение проницаемости мембраны
для калия и натрия в состоянии покоя составляет 1:0.04. Ионы калия поступают в цитоплазму и накапливаются в ней. Когда их количество достигает определенного предела, они по градиенту концентрации начинают выходить через открытые калиевые каналы из клетки. Однако уйти от наружной поверхности клеточной мембраны они не могут. Там их удерживает электрическое поле отрицательно заряженных анионов, находящихся на внутренней поверхности. Это сульфат, фосфат и нитрат анионы, анионные группы аминокислот, для которых мембрана не проницаема. Поэтому на наружной поверхности мембраны скапливаются положительно заряженные катионы калия, а на внутренней отрицательно заряженные анионы. Возникает трансмембранная разность потенциалов. Рис. Выход ионов калия из клетки происходит до тех пор, пока возникший потенциал с положительным знаком снаружи не уравновесит концентрационный градиент калия, направленный из клетки. Т.е. накопившиеся на наружной стороне мембраны ионы калия не будут отталкивать внутрь такие же ноны. Возникает определенный потенциал мембраны, уровень которого определяется проводимостью мембраны для ионов калия и натрия в состоянии покоя. В среднем, величина потенциала покоя близка к калиевому равновесному потенциалу Нернста. Например, МП нервных клеток составляет 55-70 мВ, поперечно-полосатых - 90-100 мВ. гладких мышц - 40-60 мВ, железистых клеток - 20-45 мВ. Меньшая реальная величина МП клеток, объясняется тем, что его величину уменьшают ионы натрия, для которых мембрана незначительно проницаема, и они могут входить в цитоплазму. С другой стороны, отрицательные ионы хлора, поступающие в клетку, несколько увеличивают МП.
Так как мембрана в состоянии покоя незначительно проницаема для ионов натрия, необходим механизм выведения этих ионов из клетки. Это связано с тем, что постепенное накопление натрия в клетке привело бы к нейтрализации мембранного потенциала и исчезновению возбудимости. Этот механизм называется натрио-калиевым насосом. Он обеспечивает поддержание разности концентраций калия и натрия по обе стороны мембраны. Натрио-калиевый насос —это фермент натрий-калиевая АТФ-аза. Его белковые молекулы встроены в мембрану. Он расщепляет АТФ и использует высвобождающуюся энергию для противоградиентного выведения натрия из клетки и закачивания калия в неё. За один цикл каждая молекула натрий-калиевой АТФ-азы выводит 3 иона натрия и вносит 2 иона калия т.к в клетку поступает меньше положительно заряженных ионов, чем выводится из неё, натрий-калиевая АТФ-аза. на5-10 мВ увеличивает мембранный потенциал. В мембране имеются следующие механизмы трансмембранного транспорта.
1.Активный транспорт осуществляется с помощью энергии АТФ. К этой группе транспортных систем относятся натрий-калнезый насос, кальциевый насос, хлорный насос.
2.Пассивный транспорт. Передвижение ионов осуществляется по градиенту концентрации без затрат энергии. Например, вход калия в клетку и выход из неё по калиевым каналам.
3.Сопряженный транспорт. Противоградиентный перенос ионов без затрат энергии. Например таким образом происходит натрий натриевый, натрий-кальциевый, калий -калиевый обмен ионов. Он происходит за счет разности концентрации других ионов.
Мембранный потенциал регистрируется с помощью микроэлектродного метода. Для этого через мембрану, в цитоплазму клетки вводится тонкий, диаметром менее 1 мкм стеклянный микроэлектрод. Он заполняется солевым раствором. Второй электрод помешается в жидкость, омывающую клетки. От электродов сигнал поступает на усилитель биопотенциалов, а от него на осциллограф и самописец (рис).
Дальнейшие исследования Ходжкина и Хаксли показали, что при возбуждении аксона кальмара возникает быстрое колебание мембранного потенциала, которое на экране осциллографа имело форму пика. Они назвали это колебание потенциалом действия (ПД). Так как электрический ток для возбудимых мембран является адекватным раздражителем, ПД можно вызвать, поместив на наружную поверхность мембраны отрицательный электрод - катод, а внутреннюю положительный анод. Это приведет к снижению величины заряда мембраны - ее деполяризации. При действии слабого допороговсго тока происходит пассивная деполяризация, т.е. возникает катэлектротон (.рис). Если силу тока увеличить до определенного предела, то в конце периода его воздействия на плато катэлектротона появится небольшой самопроизвольный подъём - местный или локальный ответ. Он является следствием открывают небольшой части натриевых каналов, находящихся под катодом. При токе пороговой силы МП снижается до критического уровня деполяризации (КУД), при котором начинается генерация' потенциала действия. Он находится для нейронов примерно на уровне - 50 мВ. На кривой потенциала действия выделяют следующие фазы:
1. Локальный ответ (местная деполяризация), предшествующий развитию ПД.
2. Фаза деполяризации. Во время этой фазы МП быстро уменьшается и достигает нулевого уровня. Уровень деполяризации растет выше 0. Поэтому мембрана приобретает противоположный заряд - внутри она становится положительной, а снаружи отрицательной. Явление смены заряда мембраны называется реверсией мембранного потенциала. Продолжительность этой фазы у нервных и мышечных клеток 1-2мсек.
3. Фаза реполяризации. Она начинается при достижении определенного уровня МП (примерно -20 мВ). Мембранный потенциал начинает быстро возвращаться к потенциалу покоя Длительность фазы 3-5 мсек.
4. Фаза следовой деполяризация или отрицательного следового потенциала. Период, когда возвращений МП к потенциалу покоя временно задерживается, он длится 15-30 мсек.
5. Фаза следовой гиперполяризацин или положительного следового потенциала. В эту фазу. МП на некоторое время становится выше исходного уровне ПП. Ее длительность 250-300 мсек.
Синапс (греч. synapsis соприкосновение, соединение) — специализированная зона контакта между отростками нервных клеток и другими возбудимыми и невозбудимыми клетками, обеспечивающая передачу информационного сигнала. Морфологически С. образован контактирующими мембранами двух клеток. Мембрана, принадлежащая отросткам нервных клеток, называется пресинаптической, мембрана клетки, к которой передается сигнал, — постсинаптической. В соответствии с принадлежностью постсинаптической мембраны С. подразделяют на нейросекреторные, нейромышечные и межнейрональные. Последние в зависимости от места их расположения разделяют на аксодендритические, аксосоматические, аксо-аксональные и дендро-дендритические. Наиболее сложно устроено нервно-мышечное соединение, называемое двигательной концевой пластинкой, в котором окончание аксона образует множественные синаптические контакты со специализированной мышечной мембраной. Отдельные нейроны могут иметь множество синаптических контактов с другими клетками.

Рис. 1. Синаптические контакты нейрона: синаптические бляшки (2) окончаний пресинаптических аксонов образуют соединения на дендритах (4) и соме (1) постсинаптического нейрона; 3 — аксон нейрона.
По способу передачи возбуждения с пресинаптической на постсинаптическую мембрану выделяют химические и электрические (так называемые эфапсы) синапсы. В синапсах с химической передачей возбуждения между пре- и постсинаптической мембранами имеется синаптическая щель, куда выделяется химическое вещество-передатчик — медиатор. Химические С. часто обозначают по названию медиатора (например, холинергические, адренергические, серотонинергические и т.п.). В эфапсе пре- и постсинаптические мембраны плотно соприкасаются и возбуждение передается посредством электрического тока. В зависимости от изменения биопотенциала постсинаптической мембраны различают С. деполяризующие, или возбуждающие, и гиперполяризующие, или тормозные. Механизм передачи возбуждения принципиально одинаков во всех химических С. В нем можно выделить следующие основные этапы: синтез и депонирование медиатора в пресинаптическом нейроне и его окончаниях; высвобождение медиатора из депонирующих везикул и его выход в синаптическую щель; взаимодействие медиатора со специфическими хеморецепторами постсинаптической мембраны с последующей генерацией биоэлектрического потенциала; инактивация выделенного медиатора с помощью ферментов или системы обратного поглощения. Биологически активные вещества, выполняющие функцию медиаторов, делят на несколько групп. К классическим нейромедиаторам относят ацетилхолин, адреналин и норадреналин, дофамин, серотонин, а также аминокислоты глицин и глутаминовую, аспарагиновую и гамма-аминомасляную (ГАМК) кислоты. Отдельно выделяют нейропептиды: вещество П, энкефалин, соматостатин и др. Медиаторную роль могут выполнять также АТФ, гистамин, пуриновые нуклеотиды. В соответствии с принципом Дейла, каждый отдельный нейрон в своих синаптических окончаниях один и тот же медиатор, поэтому нейроны можно обозначать также по виду медиатора: холинергические, адренергические, ГАМК-эргические, пептидергические (см. Нейросекреции). Синтез нейромедиаторов осуществляется как в соме нейрона с последующим аксонным транспортом, так и непосредственно в пресинаптических окончаниях аксона, где медиатор концентрируется в везикулах, или синаптических пузырьках.
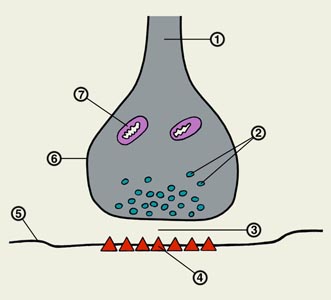
|
| Рис. 2. Межнейрональный синапс: 1 — нервное волокно (аксон); 2 — синаптические пузырьки; 3 — синаптическая щель; 4 — хеморецепторы постсинаптической мембраны; 5 — постсинаптическая мембрана; 6 — синаптическая бляшка; 7 — митохондрия. |
В состоянии функционального покоя в пресинаптическом окончании происходит случайный контакт синаптических пузырьков с пресинаптической мембраной и выделение в синаптическую щель порции (кванта) медиатора из отдельной везикулы. Например, установлено, что в С. млекопитающих квант классического медиатора ацетилхолина насчитывает 4×102—4×104 молекул. Выделившийся в синаптическую щель медиатор взаимодействует с хеморецепторами постсинаптической мембраны (рис. 2) и приводит к возникновению миниатюрного постсинаптического потенциала. Приходящий к пресинаптическому окончанию потенциал действия в несколько раз увеличивает количество выделяемого в синаптическую щель медиатора. Взаимосвязь между потенциалом действия пресинаптической мембраны и процессом выделения медиатора из везикул в синаптическую щель обеспечивается ионами Са++. Выделяющийся в синаптическую щель медиатор взаимодействует с различными хеморецептивными участками на постсинаптической мембране (см. Рецепторы). Роль мембранных, или клеточных, рецепторов играют белковые молекулы, обладающие способностью «узнавать» специфические для них вещества и вступать с ними в реакцию. В этом процессе большое значение придается системе аденилатциклаза-циклический АМФ. Предполагается, что взаимодействие медиатора с рецепторным участком аденилатциклазы приводит к образованию циклического АМФ с последующим повышением активности протеинкиназ цитоплазмы и ядра клетки. Вследствие этого интенсифицируется фосфорилирование белков, белковые молекулы подвергаются конформационным изменениям и происходит активация специальных ионных каналов мембраны. При увеличении проницаемости постсинаптической мембраны для ионов натрия, калия и хлора возникает ее деполяризация, регистрируется возбуждающий постсинаптический потенциал (ВПСП). При увеличении проницаемости лишь для ионов калия и хлора мембрана гиперполяризуется и регистрируется тормозный постсинаптический потенциал (ТПСП). ВПСП могут суммироваться, и при достижении величины деполяризации мембраны критического уровня генерируется потенциал действия. ТПСП тормозит генерацию потенциала действия, уменьшая суммарную величину ВПСП.
Основные физиологические свойства синапса обусловлены механизмом передачи возбуждения. Наличие пресинаптической мембраны с медиатором и хеморецепторов на постсинаптической мембране обеспечивает одностороннюю передачу возбуждения. Время освобождения медиатора из везикул при экзоцитозе, диффузия медиатора через синаптическую щель, взаимодействие медиатора с клеточными рецепторами постсинаптической мембраны и формирование потенциала действия создают так называемую синаптическую задержку в передаче возбуждения через С. Ее продолжительность для теплокровных животных составляет 0,2—0,5 мс. Величина синаптической задержки указывает на низкую лабильность С. по сравнению с нервными волокнами и мышцами. В связи с этим С. легко утомляется. Наличие специфических хеморецептивных участков на постсинаптической мембране делает С. высокочувствительным к биологически активным веществам. Хеморецептивные зоны часто являются точкой приложения как л СИНАПС
| — структурное образование, обеспечивающее функциональную связь нервных клеток между собой и с другими клетками (мышечными, железистыми и др.). Основной функцией нервной клетки (нейрона) является прием информации, поступающей в виде импульсов от нервных окончаний или других (промежуточных) нейронов, переработка этой информации и формирование ответных сигналов с последующей передачей их другим нервным клеткам или исполнительному органу (мышце, железистой клетке). Одна нервная клетка способна образовать до 10 000 и более связей (т. е. синапсов) на телах многих других клеток. Соответственно месту связи область контакта двигательного нервного волокна с мышечными клетками обозначают как мионевральный С, а область соприкосновения нервного волокна с железистой клеткой — как нейроэпителн-альный С. Нейроны с помощью С. объединяются в сложные комплексы или ансамбли, к-рые в свою очередь, объединяясь с помощью множества С. с другими ансамблями и группами ансамблей, образуют то единое и сложное образование, к-рое в целом называют центральной нервной системой. С. состоят из трех основных элементов: пресинаптической мембраны, пост-синаптической мембраны и синаптиче-ской щели, расположенной между ними. Пресинаптической мембраной называется мембрана нервного окончания. В пресинаптической области постоянно синтезируются физиологически активные вещества — медиаторы, заполняющие мельчайшие пузырьки. При возбуждении нейрона в нем возникает импульс, к-рый распространяется по нервному волокну и достигает пресинаптической мембраны, вызывая изменение ее физиологич. состояния. Спнап-тичес:;пе пузырьки, содержащие медиатор, лопаются, медиатор поступает в синаптическую щель. Ширина сннап-тической щели равна приблизительно 200—500 ангстрем (1 ангстрем — это одна стомиллионная часть сантиметра). Она заполнена межклеточной жидкостью, напоминающей по своему составу плазму крови. Из синаптической щели медиатор поступает к постсинап-тической мембране клетки. Постсинап-тическая мембрана чрезвычайно чувствительна к медиатору, в результате ее взаимодействия с медиатором возникает возбуждение. |
Нейрофизиологические основы психической деятельности Основой психического мира являются сознание, мышление, интеллектуальная деятельность человека, представляющие собой высшую форму адаптивного приспособительного поведения. Психическая деятельность — это качественно новый, более высокий, чем условно-рефлекторное поведение, уровень высшей нервной деятельности, свойственный человеку. В мире высших животных этот уровень представлен лишь в зачаточном виде.В развитии психического мира человека как эволюционизирующей формы отражения можно выделить следующие 2 стадии: 1) стадия элементарной сенсорной психики — отражение отдельных свойств предметов, явлений окружающего мира в форме ощущений. В отличие от ощущений восприятие — результат отражения предмета в целом и вместе с тем нечто все еще более или менее расчлененное (это начало построения своего «я» как субъекта сознания). Более совершенной формой конкретно-чувственного отражения действительности, формируемой в процессе индивидуального развития организма, является представление. Представление — образное отражение предмета или явления, проявляющееся в пространственно-временной связи составляющих его признаков и свойств. В нейрофизиологической основе представлений лежат цепи ассоциаций, сложные временные связи; 2) стадия формирования интеллекта и сознания, реализующаяся на основе возникновения целостных осмысленных образов, целостного мироощущения с пониманием своего «я» в этом мире, своей как познавательной, так и созидательной творческой деятельности. Психическая деятельность человека, наиболее полно реализующая этот высший уровень психики, определяется не только количеством и качеством впечатлений, осмысленных образов и понятий, но и существенно более высоким уровнем потребностей, выходящим за пределы чисто биологических потребностей. Человек желает уже не только «хлеба», но и «зрелищ» и соответствующим образом строит свое поведение. Его действия, поведение становятся как следствием получаемых впечатлений и порождаемых ими мыслей, так и средством активного их добывания. Соответствующим образом меняется в эволюции и соотношение объемов корковых зон, обеспечивающих сенсорные, гностические и логические функции в пользу последних.Психическая деятельность человека состоит не только в построении более сложных нервных моделей окружающего мира (основе процесса познания), но и в производстве новой информации, разных форм творчества. Несмотря на то что многие проявления психического мира человека оказываются оторванными от непосредственных стимулов, событий внешнего мира и кажутся не имеющими под собой реальных объективных причин, нет сомнения, что начальными, запускающими их факторами являются вполне детерминированные явления и предметы, отражающиеся в структурах мозга на основе универсального нейрофизиологического механизма — рефлекторной деятельности. Эта идея, высказанная И. М. Сеченовым в виде тезиса «Все акты сознательной и бессознательной деятельности человека по способу происхождения — суть рефлексы», остается общепризнанной. Субъективность психических нервных процессов заключается в том, что они являются свойством индивидуального организма, не существуют и не могут существовать вне конкретного индивидуального мозга с его периферическими нервными окончаниями и нервными центрами и не являются абсолютно точной зеркальной копией окружающего нас реального мира. Простейшим, или базисным, психическим элементом в работе мозга является ощущение. Оно служит тем элементарным актом, который, с одной стороны, связывает нашу психику непосредственно с внешним воздействием, а с другой — является элементом в более сложных психических процессах. Ощущение — это осознанная рецепция, т. е. в акте ощущения присутствует определенный элемент сознания и самосознания. Ощущение возникает как результат определенного пространственно-временного распределения паттерна возбуждения, однако для исследователей еще непреодолимым представляется переход от знания пространственно-временной картины возбужденных и заторможенных нейронов к самому ощущению как нейрофизиологической основе психики. По Л. М. Чайлахяну, переход от поддающегося полному физико-химическому анализу нейрофизиологического процесса к ощущению есть основной феномен элементарного психического акта, феномен сознания. В этом плане понятие «психическое» представляется как осознанное восприятие действительности, уникальный механизм развития процесса естественной эволюции, механизм трансформации нейрофизиологических механизмов в категории психики, сознания субъекта. Психическая деятельность человека во многом обусловлена способностью отвлекаться от реальной действительности и осуществлять переход от непосредственных чувственных восприятий к воображаемой действительности («виртуальная» реальность). Человеческая способность представить себе возможные последствия своих действий — высшая форма абстрагирования, которая недоступна животному. Ярким примером может служить поведение обезьяны в лаборатории И. П. Павлова: животное каждый раз гасило горевший на плоту огонь водой, которую оно приносило в кружке из находившегося на берегу бака, хотя плот находился в озере и со всех сторон был окружен водой. Высокий уровень абстракции в явлениях психического мира человека определяет трудности в решении кардинальной проблемы психофизиологии — нахождении нейрофизиологических коррелятов психического, механизмов превращения материального нейрофизиологического процесса в субъективный образ. Основная трудность в объяснении специфических особенностей психических процессов на основе физиологических механизмов деятельности нервной системы заключается в недоступности психических процессов прямому чувственному наблюдению, изучению. Психические процессы теснейшим образом связаны с физиологическими, но не сводятся к ним. Вторая сигнальная система. Для обеспечения несравненно более высокого уровня абстрагирования у человека появляется и развивается вторая сигнальная система: устная и письменная речь. Если даже у высших животных выработка условных рефлексов третьего и четвертого порядка представляется достаточно трудной задачей (эти рефлексы непрочны и быстро угасают), то у человека слово в виде условного обозначения, знака, не имеющего реального объективного однозначного физического содержания в виде предметов и явлений материального мира, становится достаточно сильным и прочным стимулом. Одно и то же явление, предмет на разных языках обозначаются словами, имеющими разное звучание и написание. В основе психической деятельности лежат не элементарные процессы возбуждения и торможения, а системные, объединяющие многие одновременно протекающие в мозге процессы анализа и синтеза в интегрированное целое. Психическая деятельность — функция целостного мозга, когда на основе интеграции многих нейрофизиологических механизмов мозга возникает новое качество — психика. При этом нервная модель стимула есть не что иное, как нейрофизиологическая основа формирования субъективного образа. Субъективный образ возникает на базе нервных моделей при декодировании информации и сравнении ее с реально существующим материальным объектом. В настоящее время установлены следующие достаточно определенные корреляции между различными проявлениями психической деятельности и нейрофизиологическими показателями работы мозга: 1) «волны ожидания» на ЭЭГ, которые регистрируются в ответ на сигнал, предупреждающий о предстоящей команде к действию (Г. Уолтер); 2) поздние компоненты вызванного потенциала, ассоциируемые с корковыми механизмами оценки смыслового содержания сенсорных сигналов (Л. М. Иваницкий, Э. Л. Костандов); 3) мозговые коды психической деятельности в виде определенных паттернов импульсной активности нейронов. При мульти-клеточном отведении импульсных реакций корковых нейронов установлена специфичность паттернов (узоров) импульсных потенциалов нервных клеток и нейронных ансамблей не только в отношении физических (акустических) сигналов, но и семантического (смыслового) содержания воспринимаемых слов (Н. П. Бехтерева). Психической деятельности человека эволюционно предшествуют некоторые элементы психического поведения у высших животных. К ним относится психонервная деятельность, направляемая воспроизведением образов предыдущего опыта, основанная на образном поведении животного, когда основным действенным стимулом для запускания какого-либо поведенческого акта становится не сам реальный объективный стимул окружающей среды, а «нейронный» образ этого стимула, сформировавшийся в нервных центрах (И. С. Беритов). Поведенческие акты, определяемые психонервной деятельностью, возникают при воспроизведении образа жизненно важного объекта, приводящего к удовлетворению какой-либо органической потребности животного и человека. Например, в случае индивидуального пищевого поведения таким конечным объектом является пища. Воспроизведенный «образ» пищи проецируется в определенном месте внешней среды и служит стимулом для движения животного к данному месту подобно тому, как это происходит, когда действительно пища располагается в этом месте. На определенном этапе формирования «психического» образа пищи он оказывается более сильным стимулом, чем реальная пища: животное подбегает к месту, ассоциируемому животным с пищей, но в действительности не содержащей ее (хотя животное хорошо видит, что пищи нет, но «образ пищи» оказывается сильнее реальности). Форма поведения животных и человека, определяемая образами, характеризуется тем, что при помощи проецируемых в мозге образов внешних объектов у индивида устанавливаются пространственные отношения как между этими объектами, так и между собой и ними. Психонервная активность интегрирует элементы внешней среды в одно целое переживание, производящее целостный образ. Такое воспроизведение образа может происходить и спустя длительное время после начального восприятия жизненно важной ситуации. Иногда образ может удерживаться всю жизнь без повторного его воспроизведения. Образ фиксируется в памяти и извлекается оттуда для удовлетворения господствующей биологической потребности в данный момент. В отличие от классических условных рефлексов, которые требуют повторяемости, психонервный образ формируется сразу после одной реализации поведенческого акта. Нервным субстратом, ответственным за образное отражение, очевидно, является система звездчатых нейронов с аксонами, образующими синаптические связи как с другими звездчатыми нейронами, так и через возвратные контакты с этим же звездчатым нейроном. При восприятии внешнего мира временная связь между воспринимающими сенсорную информацию звездчатыми нейронами коры большого мозга устанавливается сразу при первом одновременном или последовательном возбуждении нервных клеток, образующих проекцию данного внешнего предмета, явления. Другую форму сложных поведенческих реакций, связанных с психической сферой деятельности организма и прямо не сводимой к обычным условно-рефлекторным реакциям, представляют экстраполяционные рефлексы, основанные на способности животных и человека к прогнозированию событий, оценке, предвидению результатов своей деятельности в будущем (Л. В. Крушине кий). Экстраполяционная, или рассудочная, деятельность — это способность организма, наблюдая за течением некоторого важного события, улавливать закономерность его протекания. В результате, когда наблюдение прерывается, организм экстраполирует, т. е. мысленно продолжает ход события, соответствующим образом строя свое поведение без специальной процедуры стандартного обучения. Суть эксперимента по изучению экстраполяционной способности животного обычно сводится к следующему. Животное должно находить некий прямолинейно движущийся с постоянной скоростью объект. Особенность задачи для животного состоит в том, что первоначально видимый отрезок пути затем переходит в участок, закрытый невидимой перегородкой (ширмой), животное должно подойти к концу перегородки, учитывая, представляя себе (экстраполируя) невидимый участок, исходя из сложившейся у него в мозге картины направления движения объекта. Экстраполяционная, или рассудочная, деятельность проявляется как генетически детерминированная врожденная способность животного использовать приобретенный в течение жизни опыт в новой, незнакомой для него среде (О. С. Адрианов). Характерное свойство элементарной рассудочной деятельности заключается в способности организма улавливать простейшие эмпирические законы, связывающие предметы и явления окружающей среды, и на этой основе приобретать возможность оперировать ими при построении и реализации программ поведения в новых ситуациях. У человека эта способность развита в наибольшей степени и является одной из физиологических предпосылок, обеспечивающих возможность творческой деятельности. Экстраполяционная деятельность является важным объективным подходом к изучению элементарной рассудочной деятельности. Важнейшим элементом экстраполяции является опережение, предвосхищение будущих событий как специализированная форма отражения действительности. Возможная природа феномена опережающего отражения в структурах мозга, ответственных за высшие формы психической деятельности, по мнению П. К. Анохина, связана с разной скоростью протекания последовательных процессов в окружающей среде, природе и структурах мозга, обеспечивающих процесс отражения этой последовательности внешних явлений (рис. 15.6). Поскольку скорость процессов, протекающих в мозге, на несколько порядков выше, чем скорость процессов эволюции в окружающей среде, при достаточной длине последовательных событий на выходе системы возможно (в отражающих структурах мозга) образование модели, копии явления, предмета окружающей среды раньше, чем этот предмет, явление, событие действительно возникает в окружающем мире. Естественно, для этого надо достаточно четко и верно экстраполировать действительный ход, направление движения динамического последовательного процесса окружающей среды.
5.2. Инстинкт и другие врожденные формы поведения. Инстинкт представляет собой генетически детерминированный комплекс фиксированных действий – «наследственных координаций» по К. Лоренцу, включающих следующие компоненты: компетентное состояние особи, готовой к совершению определенного действия. Это состояние определяется нейрофизиологическими и гуморальными факторами. Так, голодное животное готово к поиску пищи (восприимчиво к соответствующим раздражителям); сексуально-восприимчивое -- к поиску партнера; ключевой раздражитель (релизер), включающий "врожденное размыкающее устройство" и тем самым, запускающий инстинктивное поведение. Так, у самца корюшки красная окраска брюшка подплывающей рыбы является ключевым раздражителем для оборонительного поведения. В состоянии готовности (компетентности) животное часто само активно ищет ключевые раздражители, а в состоянии особо сильного возбуждения инстинкт может сработать даже "вхолостую", в отсутствие релизера; завершающее (консумативное) действие, например, добывание пищи или спаривание с партнером; подавляющий раздражитель. Истинкт может не быть реализован, если имеется "мешающий" стимул, переключающий активность особи на другое поведение. Так, самец чайки может прекратить ритуал ухаживания за самкой, если он встретит пищевой раздражитель. Инстинкт часто образуют сложную иерархическую последовательность, причем завершение первого инстинктивного действия создает предпосылки для следующего действия и др. У большинства известных видов животных инстинкт сочетаются с формами поведения, приобретенными в результате обучения (см. подраздел 5.1.2), жизненного опыта. Комплексные формы поведения с инстинктивными и приобретенными компонентами (причем, вклад тех и других может варьировать), Л.В.Крушинский обозначал как "унитарные реакции". Ряд исследователей особо выделяют так называемые социальные инстинкты – врождённые механизмы, позволяющие устанавливать контакты с особями своего вида с помощью сигналов. Репертуар врожденных, генетически обусловленных форм поведения бывает столь консервативен у каждого вида, что по этим признакам виды можно классифицировать, даже если они не отличаются по морфологии. Конрад Лоренц в своих знаменитых работах 30-40х годов ХХ века продемонстрировал этот факт на примере ритуалов брачного поведения у 16 видов уток. В рамках биополитического подхода особенно важны так называемые «открытые инстинкты» (или открытые программы поведения), так что инстинктивно задана только общая канва поведения, а многие детали могут варьировать в зависимости от ситуации. Открытые программы включают танец, пение, создание семей, заботу о потомстве, охрану территории, доминирование и подчинение. Подобные открытые инстинкты, несомненно, функционируют и у человека. Помимо инстинкта, имеются и более примитивные врожденные формы поведения, не имеющие характерной для инстинкта структуры, например, кинезис (изменение скорости хаотического движения в ответ на стимул), таксис (направленное движение по направлению к стимулу или от него), рефлексы типа коленного и др. Их роль в человеческом социальном поведении, вероятно, незначительна. Более того, противники биополитического подхода как раз и пытались сводить вклад биологического элемента в поведение человеческого индивида к коленному рефлексу или отдергиванию руки от раскаленного предмета. Как мы уже видели, фактически вклад биологии существенно более весом, и о нем будет идти речь в дальнейшем тексте данного раздела. Поведение человеческих толп часто существенно примитивнее поведение индивидов. Недаром Лоренц сравнивал толпу людей в состоянии возбуждения с анонимной, безлидерной стаей рыб. Поэтому в некоторых случаях толпа обнаруживает аналоги и таких реакций как кинезис или таксис. «Ходынская катастрофа» в начале царствования Николая II и другие подобные случаи раздавливания людей неуправляемой толпой (скажем, при похоронах И.В. Сталина) в известной мере могут быть интерпретированы как хаотический «кинезис» толпы, взбудораженной сильным стимулом (раздача царских подарков, вынос тела вождя) и ведущей себя как стая рыб на нересте. КОГНИТИВНОЕ НАУЧЕНИЕ
КОГНИТИВНОЕ НАУЧЕНИЕ. Включает в себя: самоконтроль, состоящий из последовательных этапов самонаблюдения, самоподкрепления и регулирующих самооценку; составление контрактов; работу в системе правил пациента. Поведенческие правила позволяют систематизировать элементы поведения, которые можно представить следующей формулой: когда и в каких обстоятельствах делают то или другое и каковы последствия этого. Их называют еще АВС-парадигмой. Задача психотерапевта — озвучить и вербализовать эту формулу. Правила условно подразделяются так (Hoffman N., 1979):
1) правила, формулирующие ценностные установки, вызывающие определенные стимулы, которые субъективно воспринимаются по-разному, порождают у больных положительные или отрицательные эмоции (например: «Немытые овощи канцерогенны»);
2) правила, связанные с воздействием стимула (например: «После терапии все будет по-другому»);
3) поведенческие оценки (например: «Так как я заикаюсь, никто меня не слушает»);
4) правила, связанные с эмоционально-аффективным опытом личности (например: «При одном воспоминании об экзамене у меня появляется дрожь в спине», «У меня нет больше надежды»);
5) правила, связанные с воздействием реакции (например: «Я буду более пунктуальным, чтобы не вызвать гнева шефа»);
6) правила, связанные с долженствованием и возникающие в процессе социализации личности (например: «Человек должен получить высшее образование, чтобы быть счастливым»).
В процессе терапии важно установить, кто и для чего создал то или иное правило, и попытаться произвести замену негативных правил на позитивные. Большое внимание уделяется целям обучения (психотерапии). Важно работать только над теми решениями и обязательствами, которые вербализуются через «я хочу», а не «хотел бы». Формулировать проблемы лучше понятными для пациента терминами, нужно также очертить барьер, который он хочет преодолеть (например: «Я хочу преодолеть страх общения с незнакомыми людьми»). На занятии можно предложить пациенту (например, методом «мозгового штурма») найти альтернативные решения своей проблемы, записать все идеи на доске и вместе с ним выбрать самые интересные.
Пока не достигнута одна цель, переходить к другой не следует.
Психотерапевтические контракты — это запись предполагаемых изменений со стороны пациента. По возможности выбирается не бросающийся в глаза и удобный метод протоколирования и фиксируются любые изменения в процессе психотерапии.
Таким образом, при К. н., в отличие от оперантного обучения, с индивидуумом не просто что-то происходит, а то, что он сам предпринимает на основе анализа поступающей информации.
Познавательный (когнитивный) подход, обладая чертами «поведенческого», отражает суть рационально-эмоциональной психотерапии. Несовершенное мышление обнаруживается в том, что люди говорят сами себе; такой «саморазговор» воздействует и на поведение. Разновидность психотерапии, обычно включающая в себя: самоконтроль, состоящий из последовательных этапов самонаблюдения, самоподкрепления и регулирующих самооценку; составление контрактов и работу в системе правил пациента. Правила когнитивного научения условно подразделяются следующим образом (Hoffman, 1979):
- правила, формулирующие ценностные установки, вызывающие определенные стимулы, которые субъективно воспринимаются по-разному, порождают у пациентов положительные или отрицательные эмоции (например: «Немытые овощи канцерогенны»);
- правила, связанные с воздействием стимула (например: «После психотерапии все будет по-другому»);
- поведенческие оценки (например: «Так как я заикаюсь, никто меня не слушает»);
- правила, связанные с эмоционально-аффективным опытом личности (например: «При одном воспоминании об экзамене у меня появляется дрожь в спине», «У меня нет больше надежды»);
- правила, связанные с воздействием реакции (например: «Я буду более пунктуальным, чтобы не вызвать гнева шефа»);
- правила, связанные с долженствованием и возникающие в процессе социализации личности (например: «Человек должен получить высшее образование, чтобы быть счастливым»).
В процессе психотерапии важно установить, кто и для чего создал то или иное правило, и попытаться произвести замену негативных правил на позитивные. Большое внимание уделяется целям обучения (психотерапии). Важно работать только над теми решениями и обязательствами, которые вербализируются через «я хочу», а не «хотел бы». Формулировать проблемы лучше понятными для пациента терминами, нужно также очертить барьер, который он хочет преодолеть (например: «Я хочу преодолеть страх общения с незнакомыми людьми»). На занятии можно предложить пациенту (например, методом «мозгового штурма») найти альтернативные решения своей проблемы, записать все идеи на доске и вместе с ним выбрать самые интересные. Пока не достигнута одна цель, переходить к другой не следует. Психотерапевтические контракты — это запись предполагаемых изменений со стороны пациента. По возможности выбирается не бросающийся в глаза и удобный метод протоколирования и фиксируются любые изменения в процессе психотерапии. Таким образом, при когнитивном научении в отличие от оперантного обусловливания с человеком происходит именно то, что он сам предпринимает на основе анализа поступающей информации.
Поиск по сайту: