АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
Схема рефлекторного кольца
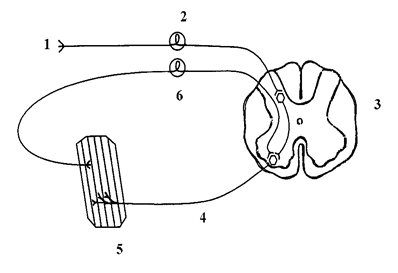
1) рецептор, 2) афферентное звено, 3) нервный центр, 4) эфферентное звено, 5) эффектор, 6) обратная афферентация.
Рефлексы отличаются большим многообразием и подразделяются на различные группы по ряду признаков.
В зависимости от расположения рецепторов выделяют экстерорецептивные и интерорецептивные рефлексы. Экстерорецептивные рефлексы вызываются раздражением рецепторов внешней поверхности тела. Интерорецептивные рефлексы могут быть висцерорецептивными и проприорецептивными. Висцерорецептивные возникают при раздражении рецепторов внутренних органов. Проприорецептивные рефлексы обусловлены раздражением рецепторов скелетных мышц, суставов, связок и сухожилий.
По характеру ответной реакции различают моторные, секреторные и сосудодвигательные рефлексы. В моторных рефлексах исполнительным органом являются мышцы. Их разновидностью являются с осудодвигательныерефлексы, которые обеспечивают изменение просвета сосудов. Секреторныерефлексы регулируют деятельность желез.
В зависимости от локализации нервных центров различают 6 основных видов рефлексов:
1) спинальные, в которых участвуют нейроны спинного мозга,
2) бульбарные, осуществляемые при обязательном участии нейронов продолговатого мозга,
3) мезэнцефальные, осуществляемые при участии нейронов среднего мозга,
4) мозжечковые, в которых принимают участие нейроны мозжечка,
5) диэнцефальные, в которых участвуют нейроны промежуточного мозга,
6) кортикальные, в осуществлении которых принимают участие нейроны коры больших полушарий.
По количеству центральных синапсов в рефлекторной дуге рефлексы подразделяются на моносинаптические и полисинаптические. Рефлекторные дуги моносинаптических рефлексов имеют два нейрона - афферентный чувствительный и эфферентный, между которыми расположен один центральный синапс. Рефлекторные дуги полисинаптических рефлексов имеют, как минимум, три нейрона: афферентный, вставочный и эфферентный.
В зависимости от продолжительности ответной реакции рефлексы могут быть:
1) фазические - быстрые и короткие,
2) тонические - продолжительные и медленные.
По биологической значимости для организма рефлексы могут быть:
1) пищевые, обеспечивающие пополнение запасов питательных веществ,
2) половые, направленные на продолжение рода,
3) оборонительные, обеспечивающие защиту организма,
4) ориентировочные, которые проявляются реакцией на новый раздражитель (рефлекс «что такое?»),
5) локомоторные, обеспечивающие движение тела.
По биологической направленности выделяют три вида рефлексов:
1) рефлексы, направленные на уравновешивание организма с внешней средой,
2) рефлексы, направленные на уравновешивание организма с внутренней средой,
3) рефлексы, направленные на продолжение рода.
И.П. Павлов выделил три основных принципа организации рефлекторных реакций организма:
1) последовательный детерминизм,
2) структурности и функции,
3) анализ и синтез.
Согласно принципу последовательного детерминизма (причинности) возбуждение по рефлекторной дуге распространяется последовательно - от рецепторов к эффекторам. При этом активация каждого последующего звена рефлекторной дуги обусловлена возбуждением предыдущего.
В соответствии с принципом структурности функции каждый морфологический элемент рефлекторной дуги выполняет специфическую функцию: рецепторы – восприятие раздражителя, афферентные нервные волокна – проведение возбуждения в ЦНС, нервный центр – анализ и синтез сигналов, эфферентные нервные волокна – проведение возбуждения к исполнительному органу.
Сущность анализа заключается в разделении поступающей в ЦНС информации на простые сенсорные сигналы. Синтез сводится к интеграции сенсорных сигналов и формированию команды для исполнительных органов. Это происходит на основе выбранной в процессе анализа наиболее важной (приоритетной) информации.
Являясь основным механизмом деятельности ЦНС, рефлексы обеспечивают поддержание гомеостаза и быстрое приспособление организма к постоянно изменяющимся условиям среды обитания. Это достигается путем сложной интеграции биоэлектрических процессов во всех отделах ЦНС.
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
Основные вопросы: Свойства нервных центров: одностороннее и замедленное проведение возбуждения, повышенная утомляемость, низкая лабильность, способность к иррадиации и суммации, последействие, трансформация ритма, пластичность, повышенная чувствительность к недостатку кислорода и питательных веществ.
Интеграция нервных процессов и рефлекторная деятельность ЦНС, лежащие в основе приспособительных реакций организма, во многом определяется общими свойствами нервных центров:
1) односторонним проведением возбуждения,
2) замедленным проведением возбуждения,
3) низкой лабильностью,
4) повышенной утомляемостью,
5) способностью к иррадиации,
6) способностью к суммации,
7) последействием (пролонгированием),
8) трансформацией ритма,
9) высокой пластичностью,
10) способность к тонической активности,
11) повышенной чувствительностью к недостатку питательных веществ и кислорода.
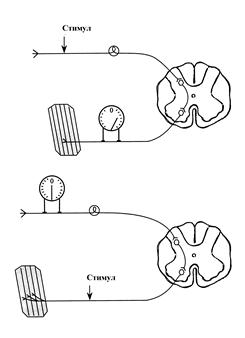
Одностороннее проведение возбуждения – это способность нервных центров проводить возбуждение только в одном направлении - от афферентов к эфферентам.
Если раздражать электрическим током афферент, то в эфферентных нервных волокнах возникает серия ПД. Однако, при раздражении эфферента, в афферентных волокнах возбуждение не возникает. Одностороннее проведение сигналов обусловлено возможностью передачи возбуждения в центральных химических синапсах только от пресинаптической мембраны к постсинаптической.
Схема эксперимента, доказывающего одностороннее проведение возбуждения в нервном центре
Замедленное проведение возбуждения - это свойство нервных центров, передавать возбуждение с низкой скоростью.
Между моментом поступления нервного импульса к окончанию аксона и формированием ВПСП на постсинаптической мембране проходит около 0,5 мс. Время от поступления нервного импульса к окончанию аксона до формирования ВПСП называется синаптической задержкой.
Низкая лабильность - это свойство нервных центров, характеризующееся относительно небольшой частотой импульсации, которую они способны воспроизвести без искажения ритма стимуляции афферента.
Лабильность нервных центров не превышает 50-70 ПД/с. Это обусловлено синаптической задержкой передачи возбуждения в центральных химических синапсах.
Повышенная утомляемость - это свойство нервных центров, которое проявляется в постепенном снижении или полном прекращении рефлекторного ответа при продолжительном раздражении афферентных волокон.
Снижение работоспособности нервных центров связано с нарушением передачи возбуждения в межнейронных химических синапсах и обусловлено повышенной утомляемостью центральных химических синапсов.
Иррадиация – это свойство нервного центра передавать возбуждение в ЦНС на другие нервные центры при достаточно сильном воздействии раздражителя на рефлексогенную зону.
Благодаря распространению возбуждения на несколько нервных центров рефлекторная реакция приобретает генерализованный характер. Иррадиации препятствуют процессы торможения, которые в нормальных условиях ограничивают распространение возбуждения в ЦНС.
В основе иррадиации возбуждения в ЦНС лежит морфологический принцип дивергенции (дихотомического ответвления аксонов нейронов), который заключается в способности клеток нервного центра устанавливать многочисленные синаптические контакты с нейронами других нервных центров
Схема иррадиации возбуждения в ЦНС
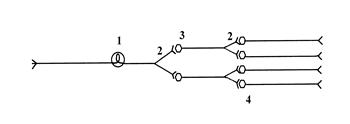
1) афферент, 2) коллатерали, 3) вставочные нейроны, 4) эфферентные нейроны.
Способность к суммации - это свойство нервных центров формировать рефлекторную реакцию при действии на рецепторы раздражителей, которые являются подпороговыми для нервного центра.
Различают два вида суммации: последовательную (временную) и одновременную (пространственную). Последовательная суммация наблюдается при взаимодействии возбуждений, которые поступают в нервный центр с коротким интервалом времени друг за другом по одним и тем же афферентам.
Если на рецептор действует один пороговый для рецептора и афферента, но подпороговый для возбуждения нервного центра раздражитель, то разрядная деятельность в области аксонного холмика центрального нейрона, а значит и рефлекторный ответ, не возникают. Однако, при последовательном действии серии таких раздражителей, в центральном синапсе увеличивается выделение медиатора. Вследствие этого на постсинаптической мембране происходит суммация ВПСП и начинается циркуляция местных ионных токов, сила которых достаточна для генерации в области аксонного холмика серии эфферентных ПД, а значит и для возникновения ответной рефлекторной реакции эффектора.
Пространственнаясуммация возникает при взаимодействии возбуждений, одновременно приходящих в нервный центр по нескольким афферентам. При одновременном действии раздражителя на несколько рецепторов одного рецептивного поля, сигналы от которых конвергирует к одному нейрону, увеличивается выделение медиатора в центральных синапсах. Вследствие этого происходит суммация ВПСП на постсинаптической мембране, и начинают циркулировать местные ионные токи, сила которых достаточна для генерации серии эфферентных ПД в области аксонного холмика, что приводит к рефлекторному ответу исполнительного органа.
Последействие - это способность нервного центра продлевать рефлекторный ответ после прекращения раздражения рецепторов.
Известны два основных механизма последействия: кратковременный и долговременный. Кратковременное рефлекторное последействие связано со следовой деполяризацией постсинаптической мембраны нейрона после прекращения раздражения афферента. После прекращения раздражения рецепторов на постсинаптической мембране центрального синапса сохраняется следовая деполяризация в виде ВПСП, который поддерживает высокую возбудимость эфферентного нейрона. В это время за счет теплового движения молекул в синаптическую щель может самопроизвольно выделиться нескольких квантов медиатора, что приводит к образованию миниатюрного ВПСП, который суммируется со следовым потенциалом. В результате этого продолжается циркуляция местных ионных токов и поддерживается разрядная деятельность эфферентного нейрона, а следовательно и рефлекторная реакция.
За счет кратковременного механизма разрядная деятельность нейрона после прекращения раздражения афферента продлевается на несколько десятков мс. По мере уменьшения амплитуды ВПСП импульсная активность нейрона постепенно уменьшается, а затем прекращается совсем.
Долговременноерефлекторноепоследействие обусловлено реверберацией - циркуляцией возбуждения по замкнутым нейронным цепям в нервных центрах.
После прекращения раздражения возбуждение продолжает циркулировать по замкнутым нейронным цепям–«ловушкам», возвращаясь к одному и тому же эфферентному нейрону. Это обеспечивает поддержание на высоком уровне его возбудимости и продолжение разрядной деятельности. Благодаря нейронным «ловушкам» возбуждение может длительно циркулировать в нервном центре до тех пор, пока не наступит утомления одного из синапсов или же активность нейронов не будет приостановлена процессом торможения.
Поиск по сайту: