АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
ВИДЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ
|
Читайте также: |
Типы мышечных сокращений. Оптимум и пессимум мышечного сокращения.
В зависимости от изменения длины мышечного волокна выделяют два типа его сокращения - изометрическое и изотоническое.
Мышечное сокращение при котором длина мышцы уменьшается по мере развиваемой ею силы, называется ауксотоническим.
Максимальная сила при ауксотонических экспериментальных условиях (с растяжимой упругой связью между мышцей и датчиком силы) называется максимумом ауксотонического сокращения. Она гораздо меньше силы, которую развивает мышца при постоянной длине, т.е. при изометрическом сокращении.
Сокращение мышцы, при котором ее волокна укорачиваются при неизменном напряжении, называется изотоническим.
Мышечная работа равна произведению расстояния (укорочения мышцы) на вес груза, который поднимает мышца. При изотонической тетанической активации мышцы от нагрузки зависит величина укорочения и скорость укорочения мышцы. Чем меньше нагрузка, тем больше укорочений в единицу времени. Ненагруженная мышца укорачивается с максимальной скоростью, которая зависит от типа мышечных волокон. Мощность мышцы равна произведению развиваемой ею силы на скорость укорочения
Сокращение мышцы, при котором ее напряжение возрастает, а длина мышечных волокон остается неизменной, называется изометрическим.
Расслабленная мышца, сохраняющая «длину покоя» за счет фиксации обоих ее концов, не развивает силу, которая передавалась бы на датчик. Но если потянуть за один ее конец, чтобы волокна растянулись, в ней возникает пассивное напряжение. Таким образом, мышца в состоянии покоя упруга. Модуль упругости покоящейся мышцы с растяжением возрастает. Эта упругость обусловлена главным образом растяжимыми структурами, которые располагаются параллельно относительно растяжимых миофибрилл («параллельная упругость»). Миофибриллы в расслабленном состоянии практически не оказывают сопротивления растяжению; актиновые и миозиновые нити, не связанные поперечными мостиками, легко скользят относительно друг друга. Степень предварительного растяжения определяет величину пассивного напряжения покоящейся мышцы и величину дополнительной силы, которую может развить мышца в случае активации при данной длине. Пиковое усилие при таких условиях называется максимумом изометрического сокращения. При сильном растяжении мышцы, сила сокращения уменьшается т. к. нити актина вытянуты из миозиновых пучков и соответственно, меньше зона перекрывания этих нитей и возможность формирования поперечных мостиков. При очень сильном растяжении мышцы, когда актиновые и миозиновые нити перестают перекрываться, миофибриллы не способны развивать силу. Это доказывает, что мышечная сила представляет собой результат взаимодействия актиновых и миозиновых филаментов(т. е. образования между ними поперечных мостиков)
В естественных условиях сокращения мышц являются смешанными - мышца обычно не только укорачивается, но изменяется и ее напряжение.
В зависимости от длительности выделяют одиночное и тетаническое сокращения мышцы.
Одиночное сокращение мышцы в эксперименте вызывают одиночным раздражением электрическим током. В изотоническом режиме одиночное сокращение начинается через короткий скрытый (латентный) период, далее следует фаза подъема (фаза укорочения), затем фаза спада (фаза расслабления) (рис. 3.1). Обычно мышца укорачивается на 5-10% исходной длины. Длительность ПД мышечных волокон также варьирует и составляет 5-10 мс с учетом замедления фазы реполяризации в конце ее.
Мышечное волокно подчиняется закону «все или ничего», т.е. отвечает на пороговое и сверхпороговое раздражение одинаковым по величине одиночным сокращением.
Сокращение целой мышцы зависит:
1. от силы раздражения при непосредственном раздражении мышцы
2. от числа нервных импульсов, поступающих к мышце при раздражении нерва.
Увеличение силы раздражения ведет к увеличению числа сокращающихся мышечных волокон.
Подобный эффект наблюдается и в естественных условиях - с увеличением числа возбужденных нервных волокон и частоты импульсов (к мышце поступает больше нервных импульсов - ПД) увеличивается число сокращающихся мышечных волокон. При одиночных сокращениях мышца утомляется незначительно.
Тетаническое сокращение - это слитное длительное сокращение скелетной мышцы. В его основе лежит явление суммации одиночных мышечных сокращений.
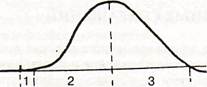
|
Рис. 6.2.1. Кривая одиночного
сокращения икроножной
мышцы лягушки:
1-латентный период,
2- фаза укорочения,
3 - фаза расслабления
При нанесении на мышечное волокно или непосредственно на мышцу двух быстро следующих друг за другом раздражений, возникающее сокращение имеет большую амплитуду и длительность. При этом нити актина и миозина дополнительно скользят друг относительно друга. В сокращение могут вовлекаться ранее не сокращавшиеся мышечные волокна, если первый стимул вызвал у них подпороговую деполяризацию, а второй увеличивает ее до критической величины. Суммация сокращений при повторном раздражении мышцы или поступлении к ней ПД возникает только в том случае, когда закончен рефрактерный период (после исчезновения ПД мышечного волокна).
При поступлении импульсов к мышце во время ее расслабления возникает зубчатый тетанус, во время укорочения - гладкий тетанус (рис. 6.2.2..). Амплитуда тетануса больше величины максимального одиночного сокращения мышцы. Напряжение, развиваемое мышечными волокнами при гладком тетанусе, обычно в 2-4 раза больше, чем при одиночном сокращении, однако мышца быстрее утомляется. Мышечные волокна не успевают восстановить энергетические ресурсы, израсходованные во время сокращения. Амплитуда гладкого тетануса увеличивается с возрастанием частоты стимуляции нерва. При некоторой (оптимальной) частоте стимуляции амплитуда гладкого тетануса наибольшая - (опти мум частоты раздражения).
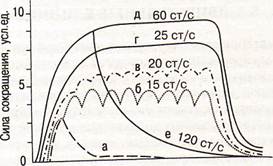
|
_Г
Раздражение нерва
Рис. Сокращения икроножной мышцы лягушки при увеличении частоты раздражения седалищного нерва (ст/с — стимулов в секунду): а — одиночное сокращение; б-д — накладывание волн сокращения друг на друга и образование разных видов тетанического сокращения. При частоте 120 ст/с - пессимальный эффект (расслабление мышцы во время стимуляции) – е.
При чрезмерно частой стимуляции нерва (более 100 имп/с) мышца расслабляется вследствие блока проведения возбуждения в нервно-мышечных синапсах - пессимум Введенского (пессимум частоты раздражения). Пессимум Введенского можно получить и при прямом, но более частом раздражении мышцы (более 200 имп/с). Пессимум Введенского не является результатом утомления мышцы или истощения медиатора в синапсе, что доказывается фактом возобновления сокращения мышцы сразу же после уменьшения частоты раздражения. Торможение развивается в нервно-мышечном синапсе при раздражении нерва.
В естественных условиях мышечные волокна сокращаются в режиме зубчатого тетануса или даже одиночных последовательных сокращений. Однако форма сокращения мышцы в целом напоминает гладкий тетанус.
Причины этого - асинхронность разрядов мотонейронов и асинхронность сократительной реакции отдельных мышечных волокон, вовлечение в сокращение большого их количества, вследствие чего мышца плавно сокращается и плавно расслабляется, может длительно находиться в сокращенном состоянии за счет чередования сокращений множества мышечных волокон. При этом мышечные волокна каждой двигательной единицы сокращаются синхронно.
Поиск по сайту: