АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ
Рефрактометрия является одним из старейших и наиболее часто употребляемых физических методов определения строения органических соединений. В настоящее время в связи с интенсивным развитием многих других современных физических методов исследования рефрактометрические методы значительно утратили свою былую ценность, но, тем не менее являются хорошими вспомогательными методами, которые помогают легко проверить правильность предположений о составе и строении органических молекул.
Рассмотрим основные физические закономерности, лежащие в основе рефрактометрического метода.
Если луч света пересекает границу раздела двух прозрачных однородных сред 1 и 2, то направление луча изменяется в соответствии с законом преломления, согласно которому, отношение синусов углов падения (i1) и преломления (i2) есть величина постоянная:

|
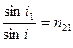
| Константа n 21 называется относительным показателем преломления второго вещества по отношению к первому. Показатель преломления вещества определяется его природой, но зависит также от внешних условий (главным образом от температуры) и от длины волны света. Длину волны указывают подстрочным индексом, а температуру − надстрочным индексом справа. Вместо длины волны |
часто употребляемых линий обычно указывают их буквенные обозначения. Например,  ,
,  и
и  обозначают показатели преломления при 20 °С для линии D натрия (5893 А) и линий С и F водорода (6563 и 4861 А).
обозначают показатели преломления при 20 °С для линии D натрия (5893 А) и линий С и F водорода (6563 и 4861 А).
Изменения плотности вещества всегда сопровождаются изменением его показателей преломления. Обычно при увеличении плотности увеличивается и показатель преломления. Существует прямо пропорциональная зависимость показателя преломления n от плотности d:
f(n) = r×d
Постоянный коэффициент r, характерный для данного вещества, называют удельной рефракцией. Величина удельной рефракции не зависит от внешних условий и агрегатного состояния вещества.
Величину удельной рефракции можно вычислить, используя формулу Лорентц-Лоренца:

Молекулярной рефракцией R называется произведение удельной рефракции на молекулярный вес М:
R = r×M
С учетом формулы Лорентц-Лоренца, получаем:

Молекулярная рефракция является мерой поляризуемости молекул. В электромагнитном поле видимого света поляризуемость молекулы практически целиком обусловлена смещением электронов и равна сумме эффектов смещений отдельных электронов. Это означает, что величина молекулярной рефракции является аддитивной. Аддитивность молекулярной рефракции органических соединений широко используется для определения их состава и строения.
Зависимость показателей преломления от длины волны называется дисперсией света или рефракционной дисперсией. В качестве меры дисперсии часто употребляется разность показателей преломления для двух длин волн nl1−nl2, называемая частной дисперсией. Частная дисперсия для спектральных линий С и F, охватывающих среднюю часть видимого спектра, называется средней дисперсией nF−nC. Для удобства эту величину часто умножают на 104 и обозначают D FC:

Родственной константой является относительная дисперсия:
 или
или 
Относительная дисперсия также обладает свойством аддитивности. В органической химии в качестве меры дисперсии используется так называемая молекулярная дисперсия, представляющая собой разность молекулярных рефракций для двух длин волн:
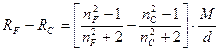
Аддитивность этой величины также позволяет использовать ее для определения строения органических соединений.
Рассмотрим применение этих величин для определения строения органических соединений.
В середине XIX в. вскоре после установления понятия о гомологическом ряде Бертло выявил простое соотношение между молекулярными рефракциями соседних членов гомологического ряда:

Согласно этому уравнению, молекулярную рефракцию n -ного члена гомологического ряда можно рассматривать как сумму молекулярных рефракций первого члена R1 и n-1 групп CH2:
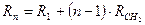
Позже данные Бертло были подтверждены большим числом примеров. Это дало основание полагать, что молекулярная рефракция, по крайней мере, для некоторых групп соединений, зависит только от природы и числа атомов в молекулах и может быть вычислена суммированием характерных для данного элемента констант − атомных рефракций:

Однако для некоторых соединений величина молекулярной рефракции, рассчитанная по данной аддитивной схеме, не совпадала с экспериментальной. Простейшими из таких соединений являлись этиленовые углеводороды и их производные. Тем не менее и этот класс соединений удалось вписать в аддитивную схему, приписав каждой двойной связи С=С особую постоянную: инкремент молекулярной рефракции двойной связи. Прибавляя этот инкремент двойной связи к сумме атомных рефракций, получали хорошее соответствие с экспериментальными значениями молекулярной рефракции. Для ацетиленовых соединений был установлен аналогичный инкремент тройной связи.
Таким образом, установление влияния природы связей на молекулярную рефракцию имело весьма большое значение, так как открывало возможность использования молекулярной рефракции для определения строения органических соединений.
В сложных функциональных группах с многовалентными элементами невозможно достаточно строго определить атомные рефракции по аддитивной схеме без условных допущений. Поэтому наряду с атомными рефракциями и инкрементами стали употребляться групповые рефракции радикалов.
В начале XX в. было выяснено, что значения молекулярной рефракции определяются главным образом числом и свойствами валентных электронов, участвующих в образовании химических связей. В связи с этим было решено видоизменить аддитивную схему и рассматривать молекулярную рефракцию как сумму рефракций связей.
Между атомными рефракциями, инкрементами и рефракциями связей существуют простые, чисто алгебраические соотношения. Например, молекулярная рефракция метана согласно аддитивной схемы равна сумме атомных рефракций или же сумме рефракций четырех связей С−Н, т.е.:

Отсюда получаем: 
При увеличении состава на группу СН2 происходит увеличение молекулярной рефракции на  и одновременное увеличение числа связей на одну связь С−С и две С−Н. Следовательно:
и одновременное увеличение числа связей на одну связь С−С и две С−Н. Следовательно:

С учетом молекулярной рефракции связи С−Н можно получить:

Таким же образом, рассматривая молекулярную рефракцию этиленовых и ацетиленовых углеводородов, можно вывести соотношения между рефракциями двойной и тройной С−С-связей и соответствующими инкрементами системы атомных рефракций:


Способ расчета по рефракциям связей более последователен, он проще и точнее передает соотношение между аддитивной схемой и современными структурными формулами. Однако при установлении строения соединений до сих пор используются как рефракции связей, так и атомные рефракции.
При установлении особых инкрементов двойной и тройной связей было замечено, что не все непредельные соединения укладываются в аддитивную схему. Особенно большие расхождения между фактическими и аддитивными значениями молекулярной рефракции наблюдались у некоторых веществ с двумя или более кратными связями С=С, СºС, С=О и т.д.
Разницу между экспериментальными и вычисленными по аддитивной схеме значениями молекулярной рефракции называют экзальтацией молекулярной рефракции и обозначают символом ЕМ:
ЕМ = Rэксп − Rадд
Величина экзальтации зависит от длины волны света, поэтому ее часто указывают подстрочным индексом.
Чаще всего величины экзальтации положительны, но в ряде случаев могут быть и отрицательными, тогда идет речь о депрессии молекулярной рефракции.
Исследование непредельных соединений показало, что особенно большие экзальтации наблюдаются при наличии систем сопряженных кратных связей. Значительные экзальтации наблюдаются также, если кратные связи находятся в сопряженном положении к бензольным кольцам, как например, в стироле, фенилацетилене и др.
Экзальтацию молекулярной рефракции обнаруживают и ароматические соединения с несколькими непосредственно соединенными или конденсированными бензольными кольцами (ряды дифенила, нафталина, антрацена и т.д.).
Молекулярная рефракция бензола и его гомологов довольно точно соответствует вычисляемой по аддитивной схеме согласно формуле Кекуле, хотя наличие трех сопряженных двойных связей в открытой цепи приводит к весьма высоким экзальтациям. Кумулированные двойные связи в алленовых и подобных им структурах также обычно не показывают экзальтаций.
Гетероциклические сопряженные системы, такие как фурановая, пиррольная и тиофеновая, обнаруживают депрессию молекулярной рефракции − значительные отклонения от аддитивности.
Одна из главных причин отклонений от аддитивности заключается в несовершенстве и упрошенном характере самой аддитивной схемы.
ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ
В конце19 столетия изучение строения и физиологии клетки привело к установлению связи ядра и обнаруженных в нем хромосом с явлениями наследственности. Редукционное деление (1 – е метотическое деление) в процессе гаметогенеза связано с распределением материнских и отцовских хромосом. Позже установили постоянство видового числа хромосом и сохранение их индивидуальности. А. Вейсман обосновал идею о том, что материальным носителем наследственности является хроматин клеточного ядра.
В 1891 – 1892 г.г. было выяснено, что перед созреванием половых клеток происходит попарное соединение (коньюгация) хромосом, которые затем расходятся при редукционном делении и эти коньюгирующие хромосомы, названные бивалентами, представляют собой пару, в которой одна хромосома материнская, а другая отцовская.
В 1902 – 1903 г.г. У. Сеттон установил связь между поведением хромосом при редукционном делении и оплодотворении и независимым расщеплением признаков в потомстве гибридов. Цитологически наблюдаемое поведение хромосом точно соответствует установленному Г. Менделем распределению наследственных факторов.
В 1905 г. Э. Вильсон сформулировал основные положения хромосомной теории определения пола. Экспериментальное доказательство локализации генов в хромосомах открыло второй период в развитии хромосомной теории наследственности, ознаменовавшейся в генетике рядом открытий.
Т. Морган в 1910 г. в лабораторных условиях проводил все свои генетические работы на плодовой мушке дрозофиле. Она маленькая и легко разводилась в пробирках на дешевом корме. У этой мушки очень короткий цикл развития и одна оплодотворенная самка дает несколько сотен мух. В соматических клетках её всего четыре пары хромосом. На основании этого с нею Т.Г. Морганом были сформулированы основные положения хромосомной теории наследственности:
1. Главными клеточными структурами, ответственными за передачу наследственной информации, являются хромосомы, которые содержат элементарные носители генетической информации.
2. В хромосомах гены расположены нитью. В пределах одной хромосомы гены образуют группу сцепления. Число групп сцепления
равно гаплоидному числу хромосом.
3. В мейозе между гомологичными хромосомами может происходить кроссинговер, благодаря которому осуществляется рекомбинация генов, что является основой биологического разнообразия видов и базой для естественного отбора организмов.
4.1 Хромосомы и наследственность
Комплекс хромосом в клетке называется хромосомным набором. Различают два основных типа хромосомных наборов : гаплоидный и диплоидный. Гаплоидный (одиночный) набор имеют клетки гаметофита (включая половые клетки) и обозначается буквой п. Диплоидный набор образуется в процессе оплодотворения и состоит из двух гаплоидных наборов – материнского и отцовского, обозначается 2п. Число хромосом для клеток одного вида – постоянно, в вегетативных (соматических) клетках может варьировать /9/. Разные виды организмов различаются между собой не только числом хромосом в наборе, но и их индивидуальной морфологией. На стадии метафазы и ранней анафазы митоза и мейоза легче всего определяются форма и размеры хромосом. В это время они имеют вид нитей или палочек, округлых в сечении. Совокупность всех морфологических признаков (включая и число хромосом) называется кариотипом, а графическое изображение кариотипа – идиограммой. Форма каждой хромосомы определяется положением первичной перетяжки, где располагается центромера. Если центромера располагается в середине, то это равноплечая или метацентрическая хромосома, если центромера слегка смещена, то это неравноплечая или субметацентрическая хромосома, если же центромера резко смещена в одну из сторон, то это резко неравноплечая или акроцентрическая хромосома. Хромосома, центромера в которой находится на конце тела называется телоцентрической. Иногда вторичная перетяжка очень заметна и отделяет от основной части небольшой фрагмент, называемый спутником.
Число хромосом посчитано для многих видов древесных растений. Морфология хромосом лучше изучена у хвойных, чем у лиственных пород.
Ген может быть определен как часть хромосомы, ответственной за проявление отдельного признака организма. Гены являются функциональными фрагментами ДНК, которые оказывают своё действие в клетке через пространство РНК и вызывают появление признака в результате длительной цепи биохимических реакций.
Гены отдельных признаков лежат в определенных местах хромосом, именуемых локусами. Хромосомы в клетках тела организма парные, поэтому каждая клетка растения (кроме половых) содержат по два гена, контролирующих один признак. Парные гены называются аллелями.
Совокупность всех генов особи составляет её генотип. Есть данные, что в клетке сосны Банкса насчитывается около 13 млн. генов. У любого вида организмов число генов всегда во много раз больше числа хромосом. Гены одной и той же хромосомы принадлежат к так называемой группе сцепления и имеют тенденцию передаваться от одного поколения к следующему как одно целое. Факт сцепления генов необходимо учитывать при селекции.
Существует классификация генов, основанная на характере их действия (Дж. Райт, 1978).
Аддитивные гены имеют слабый эффект, контролирующий один и тот же признак, взаимно усиливают этот эффект кумулятивным образом.
Гены доминантные и рецессивные. Доминантный ген (А, В, С…) вызывает выраженность признака. Рецессивный ген (а, в, с …) проявляется слабо или совсем не проявляется, кроме тех случаев, когда он присутствует в обеих парных хромосомах, т.е. унаследован от обоих родителей. Внешнее
проявление наследственных признаков (генотипа) называется фенотипом.
Гены эпистатические, т.е. они доминантны по отношению к другому гену (гипостатичному).
Гены сверхдоминантности, если гетерозигота Аа способствует выраженности признака более полно, чем каждая из гомозигот (Аа и аа) родителей. Сверхдоминантность – одно из возможных объяснений гибридной мощности (превосходства гибридов над каждым из родителей).
Взаимное расположение генов в хромосоме можно установить путём скрещивания так называемых чистых линий, у которых известные анализируемые признаки находятся в гомозиготном состоянии. При мейозе гомологичные хромосомы могут обрываться и обмениваться частями. Если это произошло, то говорят, что имел место кроссинговер.
4.2 Определение развития пола
Каждому виду свойственно определенное соотношение полов. Чаще оно равно 1:1. Хромосомы, составляющие одну пару в хромосомном наборе мужской и женской особей, по форме и величине очень похожи, кроме одной пары, в которой хромосомы значительно отличаются друг от друга. Установлено, что с этими необычными хромосомами связано определение пола и их называют половыми хромосомами. В хромосомном наборе животных и раздельнополых растений различают обычные хромосомы или аутосомы и половые X и Y – хромосомы. По хромосомной теории наследственности пол организма определяется в момент оплодотворения.
У многих видов особи женского пола имеют парные хромосомы, и в результате редукционного деления у них образуется только один сорт гамет X, а в гаметогенезе мужского пола образуется два сорта гамет : X и О либо X и Y при равном числе аутосом. Пол, дающий гаметы одного сорта - X X – называется гомогаметным, дающий два сорта гамет X и О и X и Y – гетерогаметным. Следовательно, если самка определяется наличием в зиготе двух Х – хромосом, а самец – наличием лишь одной Х – хромосомы и мужские гаметы с Х и О образуются в равном числе, то в потомстве окажется соотношение полов.
Р ♀ХХ х ♂ХО ♀ХХ х ♂ХY
F1 1♀XX х 1♂XO или 1♀ХХ: 1♂XY
Это явилось первым научным объяснением расщепления по полу 1:1.
Существует четыре основных типа хромосомного определения пола (таблица 1).
Таблица 1 – Типы соотношения половых хромосом у животных и растений
| Организмы | Гетерогаметный пол | Гаметы | Зиготы | ||
| яйцеклетки | спермии | самки | самцы | ||
| Человек, дрозофила, древесные виды (раздельнополые) | мужской | Х и Х | Х и Y | Х Х | Х Y |
| Клоп | мужской | Х и Х | Х и О | Х Х | Х О |
| Кузнечик | мужской | Х и Х | Х и О | Х Х | Х О |
| Птицы, бабочки | женский | Х и Y | Х и Х | Х Y | Х Х |
| Моль | женский | Х и О | Х и Х | Х О | Х Х |
4.3 Пол и половые хромосомы у растений
Подавляющее большинство высших растений гермафродитные. Гаметы таких организмов генетически совершенно идентичны. Но около 5% цветковых растений двудомные: тычинковые и пестичные цветки у них находятся на различных особях (женских и мужских). У таких организмов выражен половой диморфизм. Среди двудомных видов имеются ценные сельскохозяйственные растения: виноград, конопля, дынное дерево и др. Зависимость между полом и структурой хромосом у таких растений была обнаружена позднее, чем у животных. Вначале установили сцепленное с полом наследование у дрёмы белой, а затем у этого растения, а также у щавеля, хмеля и др., были открыты половые хромосомы. У растений существует два основных типа генетического контроля пола (таблица 2).
Таблица 2 – Хромосомный механизм определения пола у двудомных растений
| Вид | Число хромосом (2п) | Пол | |
| женский | мужской | ||
| Конопля посевная | ХХ | ХY | |
| Щавель малый | ХХ | ХY | |
| Спаржа | ХХ | ХY или YY | |
| Дрема белая | ХХ | ХY |
Первый из них определяется присутствием или отсутствием Х – хромосомы. Второй тип связан с факторами, находящимися в половых хромосомах и аутосомах, и развитие признаков того или иного пола определяется соотношением половых хромосом и аутосом.
4.4 Наследование признаков, сцепленных с полом
Признаки, определяемые генами, находящимися в Х- хромосомах, называются признаками, сцепленными с полом. Пол любого организма определяется генетическим механизмом, в основе которого лежит комбинация половых хромосом. Распределение в мейозе признаков, гены которых находятся в половых хромосомах, должно соответствовать поведению половых хромосом.
Совместное (сопряженное) наследование неаллельных генов, расположенных в паре гомологичных хромосом, называется сцепленным. Если все гены находятся в одной хромосоме, то говорят о группе сцепления.
Число групп сцепления у организма соответствует числу хромосом в гаплоидном наборе.
Генетическими исследованиями установлено, что у дрозофилы Y – хромосома, в отличие от Х – хромосомы, не содержит генов, т.е. наследственно инертны.
Поэтому гены, находящиеся в Х – хромосоме, как правило, не имеют аллелей в Y – хромосоме. В силу этого рецессивные гены в Х – хромосоме гетерогаметного пола могут проявляться, будучи в единственном числе. Следовательно, признаки, гены которых находятся в половых хромосомах, должны наследоваться по принципу, отличающемуся от менделевского расщепления при доминировании. Признаки, определяемые генами, находящимися в Х – хромосоме, наследуются сцепленно.
Тип скрещивания, когда гены матери наследует сын, а гены отца – дочь, называется крисс – кросс (крест – накрест).
МОЛЕКУЛЯРНЫЕ ОСНОВЫ НАСЛЕДСТВЕННОСТИ
Цитологический анализ показал, что материальной основой наследственности являются хромосомы. Отдельные участки хромосом – локусы определяют развитие различных признаков и свойств организма.
Хромосомы состоят из белка гистона (или иротамина) и нуклеопротеидов (до – 92%). Нуклеопротеиды представлены дезоксирибонуклеиновой кислотой (ДНК). В хромосомах присутствует рибонуклеиновая кислота (РНК), некоторые количества ионов кальция, магния, железа и других элементов, и негистонные белки, иногда образующие комплекс с РНК.
5.1. ДНК и РНК
Нуклеиновые кислоты представляют собой линейные полимеры, очень крупные молекулы, которые образованы цепочкой звеньев - нуклеотидов, каждый из которых состоит из трех компонентов: азотистого основания, остатка сахара и остатка фосфорной кислоты. ДНК содержит сахар - дезоксирибозу, а РНК – рибозу. Оба эти сахара принадлежат к группе пентоз. В состав ДНК входят четыре типа азотистых оснований: пуриновый – аденин (А) и гуанин (Г) и пиримидин – цитозин (Ц) и тимин (Т). РНК тоже имеет четыре типа основания, из которых три (аденин, гуанин и цитозин) такие же как у ДНК, а тимин заменен другим пиримидином – урацилом (У).
Связь между нуклеотидами в цепи ДНК строго однотипна и осуществляется за счет образования диэфира фосфорной кислоты между определёнными (третьим и пятым) гидроксилами соседних дезоксирибозных остатков. К каждому дезоксирибозному остатку этой каркасной цепи присоединены боковые радикалы – пуриновое или пиримидиновое основание.
Последовательность нуклеидов в макромолекуле ДНК у разных видов не одинакова, это имеет прямое отношение к наследственности. Отношение количества молей (гуанин + цитозин) к количеству молей (аденин + тимин) у разных видов колеблется от 0,45 – 2,8. В одной молекуле ДНК объединяются две полинуклеотидные цепочки в виде двойной спирали с правым ходом винта, диаметром около 20А (ангстрем). При этом пуриновые и пиримидиновые основания обеих цепей оказываются заключенными внутри пространства между витками спирали. Азотистые основания обеих нитей ориентированы по направлению к середине спирали, причем аденин одной нити всегда находится напротив тимина другой нити, а гуанин первой нити напротив цитозина другой нити. Каждое из этих оснований соединены друг с другом водородными связями, две такие связи имеются в паре – аденин – тимин и три в паре гуанин – цитозин. Следовательно, обе нити ДНК взаимно дополняют друг друга, что называется комплиментарностью.
В клетках организма одного вида количество гуанина равно количеству цитозина, а количество аденина – тимину, суммарное количество А+Г=Т+Ц. Отношение Г+Ц к А+Т специфично для каждого вида, даже для деревьев разного пола (у осин).
Важным свойством ДНК является её способность к самоудвоению (редупликации, ауторепродукции), благодаря чему сохраняется постоянство ДНК в ряду клеточных поколений. В определенный момент жизни клетки в молекулах ДНК разрываются водородные связи и цепи расходятся. Далее каждая цепь строит вторую недостающую по уже известному принципу комплиментарности.
В процессе синтеза новой нити участвуют ферменты, называемые ДНК – полимеразами. Они производят репарацию (залечивание мелких повреждений, которые бывают при репликации), инициацию (начало синтеза каждого нового фрагмента), элонгацию (наращивание синтезируемой новой нити) и терминацию (окончание синтеза каждого нового фрагмента).
В отличие от ДНК молекула РНК состоит из одной длинной и разветвленной цепи. Известны три вида РНК: информационная (и – РНК), называемая иногда матричной РНК, транспортная (т – РНК) и рибосомная (р – РНК). Молекула и – РНК состоит из сотен нуклеинов, длина её составляет от тысячи до нескольких тысяч ангстрем; и – РНК передаёт наследственную информацию из ядра цитоплазмы; т – РНК представлена двадцатью различными формами – по числу аминокислот, входящих в состав молекул белков и состоит, примерно, из семидесяти нуклеотидов. С помощью т- РНК аминокислоты доставляются к месту синтеза белка – рибосомами. Рибосомная РНК входит в состав рибосом клетки.
В результате взаимодействия трёх указанных типов РНК в клетке происходит синтез специфических ферментов и всех белков. Процесс связывания отдельных нуклеотидов в молекулы нуклеиновых кислот называется полимеризацией.
5.2 Структура гена, генетический код
Участок молекулы состоит из нескольких нуклеотидов и контролирующих формирований элементарного признака через синтез белков, называемого геном.
Весь генетический материал разделяется на чистроны – единицы функции, мутоны – единицы мутации и риконы – единицы рекомбинации. Все эти единицы характеризуются разным количеством пар нуклеотидов.
Гены делятся на две категории:
1) Структурные, координирующие строение определённых белков;
2) Функциональные (акцепторные), служащие местом специфического присоединения белков – репрессоров и белков – активаторов.
К акцепторным генам относится: ген – оператор, ген – промотор, ген- терминатор. Ген – оператор координирует проявление соседних генов, составляющих опером. Опером – это функционально генетическая единица, размером средним между размерами гена и хромосомы. Он составлен из ряда линейно расположенных генов.
Ген – промотор это стартовые точки на ДНК, к которым присоединяются РНК – полимеразы, чтобы начать транскрипцию. Транскрипция – это переписывание последовательности нуклеотидов гена с ДНК на РНК.
Трансляция – это перевод последовательности нуклеотидов гена в последовательность аминокислот белка.
Ген – терминатор – это ген, прекращающий определенные действия других генов.
Последовательность нуклеотидов ДНК, составляющих ген, кодирует последовательность аминокислот в белке. Всего известно двадцать основных аминокислот. Четыре типа нуклеотидов – А, Г, Ц, Т, сгруппированные по три (например, ЦЦТ, ГУА, УУЦ т.д.), могут кодировать 64 аминокислоты (43=64).
Три нуклеотида, кодирующие одну аминокислоту в ДНК, называются кодоногеном, а в и – РНК – кодоном. Каждый кодоноген присоединяет комплиментарный кодон. В процессе трансляции в рибосомах к кодонам
и – РНК подходят транспортные РНК (т – РНК) соответствующими антикодонами.
В результате исследований Ф. Крика, Г. Кораны, М. Ниренберга и др. в 1965 году был составлен генетический код в его современном виде. При этом отмечаются следующие его особенности:
1. Код является триплетным, т.е. каждая аминокислота кодируется
группой из трех нуклеотидов (триплетом нуклеотидов).
2. Код является неперекрывающимся.
3. Код вырожден, т.е. одна аминокислота может кодироваться
несколькими определенными триплетами нуклеотидов.
4. Код не имеет никаких "запятых", т.е. кодоны ничем не отделены
друг от друга.
5. Код считывается с фиксированной точки в пределах гена,(моле-
кулы нуклеиновой кислоты) в одном направлении.
Аминокислота, принесенная т – РНК, только в том случае идет на синтез белка, когда триплет РНК комплиментарен триплету т – РНК.
5.3 Основные типы мутаций и принципы их классификации
Мутации – внезапные естественные или вызванные искусственно, наследуемые изменения генетического материала, приводящие к изменению этих или иных признаков организма.
Мутации классифицируют по различным признакам, в зависимости от того, на каком уровне они возникают и какие генетические структуры затрагивают.
Классификация мутаций по характеру изменения генома:
а) генные мутации – изменение генов;
б) хромосомные мутации или хромосомные перестройки – изменение структуры хромосом;
в) геномные мутации – изменение числа хромосом.
Генные мутации представляют собой изменение чередования пар нуклеотидов в ДНК, которые составляют класс точковых или точечных мутаций. Этот класс мутаций подразделяется на следующие группы: транзиции, трансверсии, вставки, выпадания (рисунок 2).
 Хромосомные перестройки
Хромосомные перестройки









 Внутрихромосомные Межхромосомные
Внутрихромосомные Межхромосомные
 |  |  |  | ||||||||
|
| ||||||||||
Рисунок 2 – Схема хромосомных перестроек
(С.Г. Инге – Вечтомов, 1989г.)
Хромосомные мутации возникают в следствии разрывов хромосом.
Межхромосомные перестройки: транслокации – перемещение части хромосомы на другую хромосому, гомологичную ей; транспозиция – изменения локализации небольших участков генетического материала, включающих один или несколько генов.
Элементарной единицей мутации считается мутон, равный одному нуклеотиду.
1. По проявлению в гетерозиготе:
а) доминантная мутация;
б) рецессивная мутация;
2. По уклонению от нормы и так называемого дикого типа:
а) прямые мутации;
б) реверсивные мутации, включающие как обратную, так и
супрессорные мутации.
3. В зависимости от причин, вызывающих мутации:
а) спонтанные - возникающие без единой причины;
б) индуцированные – вызванные искусственно.
4. По локализации в клетке:
а) ядерные;
б) цитоплазматические, т.е. мутации неядерных генов.
5. По характеру фенотипического проявления:
а) морфологические;
б) физиологические;
в) биохимические.
6. По отношению к возможностям наследования:
а) генеративные – происходящие в половых клетках;
б) соматические, происходящие в соматических клетках.
Помимо вышеперечисленных существуют более частные принципы классификации мутаций – летальные, малые, обратные, пластидные.
Если изменения пропорциональные (или кратные) гаплоидному набору хромосом, то говорят о полиплоидии; если изменяется число хромосом внутри гаплоидного набора – об анеуплоидии.
Факторы, вызывающие изменение в молекулах ДНК или хромосомах, называются мутагенами (ионизирующие радиации, ультрафиолетовые лучи, температурные шоки, лазерное излучение, а также многие химические соединения).
ЗАКОНОМЕРНОСТИ НАСЛЕДОВАНИЯ ПРИЗНАКОВ ПРИ ВНУТРИВИДОВОЙ ГИБРИДИЗАЦИИ
Анализ особенностей наследования заключается в изучении степени сходства в изучении степени сходства или различия в проявлении признаков у родителей и потомства.
Процесс естественного или искусственного соединения двух генетических разнородных гамет при оплодотворении называется скрещиваниемили гибридизацией, потомство получаемое при этом, называется гибридным. Внутривидовое скрещивание между особями, различающимися по одному гену, называется моногибридным, по двум генам – дигибридным, по трем - тригибридным и т.д.
6.1 Метод генетического анализа
Закономерности наследования впервые были сформулированы и доказаны экспериментально Г. Менделем. Многочисленные попытки установления закономерности наследования до Г. Менделя оставались безуспешными. Причина этого заключается частично в неудачном подборе объектов для исследования, а главным образом – в ошибках применяемой методики. Объекты брались без учета особенностей их опыления, порой без проверки устойчивости изучаемых признаков.
Г. Мендель проводил опыты с горохом – самоопыляющейся культурой с нормальной плодовитостью и наличием многочисленных форм по различным признакам.
Под формой подразумевается совокупность особей одного вида, отличающихся от других особей того же вида внешним проявлением одного или нескольких признаков. Менделем была разработана оригинальная методика исследования, заключающаяся в количественном учете гибридных растений с применением индивидуального анализа признаков в потомстве от каждого материнского растения в ряду поколений. Родители подбирались так, чтобы они отличались внешним проявлением какого – то одного признака, например, окраской цветов или семенной кожуры, высотой растений, формой семян и т.д. Перед скрещиванием родительские растения самоопылялись, и в течении нескольких поколений изучался характер проявления интересующего признака в потомстве. Обязательным условием опыта являлась устойчивость признака у родителей при самоопылении. Скрестив подобранные пары, Г. Мендель подсчитывал количество растений с одним и другим выражением признака в потомстве от каждой пары и суммарно. Гибриды первого поколения он заставлял самоопыляться и в F2 так же учитывал количество растений с различным выражением признака от каждого исходного растения отдельно. Так он получал растения до F7. Тщательно продуманный выбор объекта и оригинальная методика позволили
Г. Менделю установить общие закономерности наследования при внутривидовой гибридизации.
6.2 Наследование при моногибридном скрещивании. Законы Г. Менделя
В результате скрещивания растений или животных, имеющих по тем или иным признакам наследственные различия, получаются гибридные организмы или гибриды. Скрещивания, в которых родительские формы отличаются по одной паре альтернативных признаков, называются моногибридными. Изучение явлений наследственности Г.Мендель начал с простейших моногибридных скрещиваний, а затем проводил гибридизацию сортов, различающихся по двум и большему числу признаков.
При опылении гороха с красными цветками пыльцой, взятой с растений гороха с белыми цветками, все гибриды первого поколения были с красными цветками. Такие же результаты получены при обратном скрещивании. Красная окраска цветков неизменно сохранялась, белая же подавлялась и не проявлялась. Признак, сохраняющийся у гибридов первого поколения Г.Мендель назвал доминантным (от dominantis – господствующий, подавляющий), признак, непроявляющийся – рецессивным (от латинского recessives – отсутствующий, подавляемый). Следовательно, наследуются не признаки, а их потенциальные возможности, которые Г.Мендель назвал факторами (в современном понимании это гены). Факторы наследуются через гаметы, и у гибридных растений не смешиваются друг с другом, а относительно независимы друг от друга. Явление наследования в гаметах гибридного организма наследственных факторов Г.Мендель назвал правило чистоты гамет.
Подавление у гибридных организмов одних признаков другими получило в генетике название доминирования. Почти во всех опытах, которые проводил Г.Мендель, доминантный признак полностью подавлял проявление рецессивного признака, поэтому гибриды F1 были одинаковыми между собой и с родительскими растениями, имеющими доминантный признак. На основании этого Г.Мендель сформулировал правило единообразия гибридов первого поколения.
Продолжая опыты, Г.Мендель собрал семена гибридов F1 отдельно с каждого растения и высеял их. Особи второго поколения не были сходны между собой. Так растения F2, выращенные из семян красноцветковых гибридов первого поколения, имели как красные, так и белые цветки. Подсчеты показали, что на три красноцветковых растения приходилось одно белоцветковое (точное соотношение: 3,01 красноцветкового на одно белоцветковое). Это отношение не представляло исключения. Оно наблюдалось по всем другим парам признаков, участвующих в скрещивании. Этот фактор, показавший, что рецессивный признак в скрытом виде проходит поколение гибридов и вновь возникает, как говорят, выщепляется в потомках гибридов, привел Г. Менделя к идее о существовании ответственных за эти явления наследственных факторов. Он обозначил наследственный фактор для доминантного признака буквой А и наследственный фактор для рецессивного признака буквой а. Гибриды обладали факторами А и а, поэтому их структура записана как Аа. Чистые доминантные особи – АА, а рецессивные аа. В дальнейшем организмы, имеющие в паре символов одинаковые факторы АА и аа, стали называть гомозиготами, а особи – Аа получили название гетерозигот. Закономерность в распределении доминантных и рецессивных признаков у гибридов второго поколения в кратном отношении 3:1 Г. Мендель назвал – правилом расщепления гибридов второго поколения.
С целью разграничения внешних признаков и наследственных задатков В. Иоганнсен в 1903 году предложил термины фенотип и генотип.
Под фенотипом понимается совокупность внешних признаков организма, а под генотипом– совокупность его генов. В данном случае фенотип – это окраска цветков, а генотип – наследственные задатки (гены), обозначенные буквами А и а, контролирующие формирование фенотипических признаков.
Развитие цитологических методов исследования подтвердило правильность гипотезы Г. Менделя, показав силу его научного предвидения существования генов и механизма их распределения по гаметам (мейоза).
6.3 Наследование при дигибридном и полигибридном скрещиваниях
Скрещивания, в которых родительские формы отличаются по двум парам признаков, называются дигибридными, а если число признаков больше – полигибридными.
Для анализа при дигибридном скрещивании были окраска и характер поверхности семян гороха. Жёлтая окраска А доминировала над зелёной а, а гладкая поверхность В – над морщинистой в. Гены А и а являются одной аллельной парой, Вв – второй. Между собой две пары генов А и а,В и в неаллельны. Принципиальное отличие от моногибридного скрещивания заключается в том, что в мейозе учитывается не одна, а две пары гомологических хромосом. Одна из хромосом каждой пары может отойти как к одному, так и к другому полюсу клетки, вторая хромосома из гомологической пары выбора не имеет, а отходит только к противоположному полюсу.
Гетерозиготные растения по обеим парам (F1) образуют четыре типа гамет и при дигибридном скрещивании, как и при моногибридном, все растения F1 получаются одинаковыми и по генотипу (дигетерозиготные), а по фенотипу (с доминирующими признаками).Для расчёта сочетаний разных типов гамет и определения результатов расщепления пользуются решёткой Р. Пеннета, названной по имени английского генетика, предложившего этот приём.
По вертикальной линии, расчерченной в клетку, таблицы наносят типы яйцеклеток, а по горизонтальной – типы спермиев. На пересечение линий, ведущих от обозначений обоих типов гамет, выписывают сначала гены одной аллельной пары, а затем другой. Так определяют генотипы и соответствующие им фенотипы гибридов при всех возможных сочетаниях яйцеклеток и спермиев.
 |
| ♀ ♂ | Г а м е т ы | ||||
| АВ | Ав | аВ | ав | ||
| Г а м е т ы | АВ | ААВВ | ААВв | АаВВ | АаВв |
| Ав | ААВв | ААвв | АаВв | Аавв | |
| аВ | АаВВ | АаВв | ааВВ | ааВв | |
| ав | АаВв | Аавв | ааВв | аавв |
Решетка Пеннета позволяет определить все возможные сочетания мужских и женских гамет и частоту их встречаемости.
В F2 расщепление более сложное. Образуются 9 генотипических классов растений в соотношении 1:2:2:4:1:2:1:2:1 и четыре фенотипических класса в соотношении 9:3:3:1. В этом заключается сущность второго правила Г. Менделя при дигибридном скрещивании. Если у растений наблюдались два сочетания признаков (жёлтые гладкие и зелёные морщинистые), то в F2, кроме этих сочетаний, получились растения с новым сочетанием признаков (жёлтые морщинистые и зелёные гладкие), то есть были реализованы все четыре сочетания, возможные по правилам свободного комбинирования. На этом основании Г.Мендель сформулировал третье правило независимого комбинирования генов.
Полигибридные скрещивания дают более сложную картину расщепления по сравнению с дигибридными скрещиваниями, но подчиняются тем же законам наследования.
Число возможных комбинаций гамет и число классов по фенотипу и генотипу можно определить не составляя решетки Пеннета, а пользуясь таблицей 3.
Таблица 3 – Число классов гибридных особей по фенотипу и генотипу и ха-
рактер расщепления в F2 при различном числе пар признаков (полное доминирование).
| Скрещи- вание | Число пар приз-наков | Число образующихся гамет | Число возмож- ных комбинаций гамет | Число классов | Числовое от- ношение классов по фено- типу | |
| по фенотипу | по генотипу | |||||
| Моногибридное | 21=2 | 41=4 | 21=2 | 31=3 | 3:1 | |
| Дигибридное | 22=4 | 42=16 | 22=4 | 32=9 | 9:3:3:1 | |
| Тригибридное | 23=8 | 43=64 | 23=8 | 33=27 | 29:9:9:9:3:3:3:1 | |
| Тетрагибридное | 24=16 | 44=256 | 24=16 | 34=81 | 81:27:27:27:27 :9:9:9:9:9:9:3:3 :3:3:1 | |
| Полигибридное | n | 2n | 4n | 2n | 3n | (3:1)n |
Для этого должно быть известно, по скольким парам аллеломорфных признаков различаются скрещиваемые формы. Соединение гамет с различными генами во время оплодотворения происходит совершенно случайно и подчиняется законам теории вероятности. Чем больше гибридных особей, тем сильнее фактически полученные данные будут приближаться к теореотчески ожидаемым.
Ряды расщепления представляют собой бином (3+1)n, где n – количество пар аллелей, по которым наблюдается расщепление.
Закономерное расщепление при полигибридном скрещивании возможно при соблюдении двух дополнительных условий: нахождение учитываемых генов в негомологичных хромосомах; равновероятном образовании всех сортов гамет на основе случайного расхождения негомологичных хромосом в мейозе.
НАСЛЕДОВАНИЕ ПРИ ВЗАИМОДЕЙСТВИИ ГЕНОВ
7.1 Аллельное и неаллельное взаимодействие генов
Правильность установленных Г. Менделем закономерностей наследственности была подтверждена после 1900 года в многочисленных опытах по изучению наследования различных признаков как у растений, так и у животных. Выяснилось, что полученные Г. Менделем определённые числовые отношения при расщеплении в потомстве гибридов были верными во всех тех случаях, когда каждый ген определял развитие одного наследственного признака. Выяснилось, что:
1. Один и тот же ген может влиять на несколько различных признаков;
2. Происходит взаимодействие генов, когда один и тот же наслед-
ственный признак развивается под влиянием многих генов.
Таким образом, фенотипическое выражение большинства признаков и свойств организма определяется в онтогенезе взаимодействием многих генов. Это отражается и на характере расщепления гибридов разных скрещиваний, особенно если родительские формы отличаются по нескольким признакам.
Влияние одного гена на развитие двух и большего числа признаков называется множественным или плейотропным действием, а само явление – плейотропией. Оно очень широко распространено: большинство генов у всех организмов действует плейотропно. Примеров плейотропного действия генов много. Замечено, что у растений с пурпурными цветками всегда имеются красные пятна в пазухах листьев, а семенная кожура серого или бурого цвета. Все эти три признака определяются действием одного наследственного фактора.
Известны два вида взаимодействия генов: аллельное и неаллельное. Простейший пример аллельного взаимодействия генов – неполное доминирование при скрещивании красноцветковых и белоцветковых растений львиного зева. Розовая окраска цветка у гибридов F1 в этом скрещивании – результат взаимодействия двух аллельных генов А и а. Полное доминирование также результат взаимодействия двух генов одной аллельной пары. Взаимодействие генов имеет биохимическую природу и основано на взаимодействии синтезируемых под контролем генов белков ферментов.
Взаимодействие неаллельных генов проявляется в четырёх основных формах: комплиментарности, эпистазе, полимерии и модифицирующего действия генов. Каждая из этих форм приводит к характерным изменениям известных числовых отношений при расщеплении в дигибридных скрещиваниях.
7.2 Типы взаимодействия неаллельных генов
Комплиментарное (дополнительное) действие генов наблюдается в случаях, когда неаллельные гены раздельно не проявляются, но при одновременном присутствии в генотипе обуславливают развитие нового признака. При этом признак развивается в результате взаимодействия двух ферментов, образуемых под контролем двух неаллельных генов. Например, при скрещивании двух форм душистого горошка, имевших белые цветки, все гибридные растения в F1 оказались с красными цветками. При самоопылении этих растений в F2 признаки расщепились в отношении: 9 красноцветковых: 7 белоцветковых.
 Подавление действия одной аллельной пары генов доминантным геном другой, неаллельной им пары генов называется эпистазом. Явление эпистаза можно выразить формулой
Подавление действия одной аллельной пары генов доминантным геном другой, неаллельной им пары генов называется эпистазом. Явление эпистаза можно выразить формулой  , когда один доминантный ген не допускает проявления другого доминантного гена.
, когда один доминантный ген не допускает проявления другого доминантного гена.
Гены, подавляющие действие других, не аллельных им генов, называются эпистатичными, а подавляемые – гипостатичными. Проявление эпистаза наследования окраски плодов можно проследить на примере тыквы. У этого растения известны 3 типа окраски плодов: белая, жёлтая и зелёная. При скрещивании растений с белыми и жёлтыми, а также белыми и зелёными плодами доминирует белая окраска, т.к. у тыквы окраска плода контролируется двумя парами генов. При этом, очевидно имеется ген белого цвета W, эпистатичный по отношению к генам жёлтой и зелёной окраски плодов. Он подавляет образование любого другого пигмента, и в его присутствии, независимо от наличия или отсутствия других генов никакая окраска проявиться не может.
Особый интерес для селекционеров представляет случай, когда развитие того или иного признака зависит от двух или большего числа одинаково действующих генов.
Неаллельные гены, действующие на формирование одного и того же признака, называются полимерными, или множественными. Явление взаимодействия неаллельных множественных генов, обуславливающих его развитие, называется полимерией. При полимерии два или несколько ферментов, образующихся под контролем неаллельных генов, действуют на развитие одного признака, усиливая его проявление. Простейшим примером полимерии является наследование двух типов окраски зерна. У пшеницы: краснозерный, имеющий в оболочке зерновых красный пигмент и белозерный, лишенный его. Признак красной окраски доминирует над белой. Как правило, при скрещивании краснозерных сортов с белозерными в F2 идёт расщепление по обычной моногибридной схеме: 3 краснозерных: 1 белозерное. Но при скрещивании некоторых сортов пшеницы, имеющих темноокрашенное зерно, с белозерными сортами, когда растения в F1 также дают окрашенное зерно, в F2 происходит в отношении: 15 краснозерных: 1 белозерное растение. Но интенсивность окраски зерна у краснозерных растений различна. Она варьирует от темно – до бледно – красной, т.е. интенсивность окраски зависит от количества доминантных генов, действующих на этот признак, примерно, в равной степени.
При изучении взаимодействия были открыты гены основного действия, т.е. такие, которые определяют развитие признака или свойства, например, выработку пигментов, форму цветка и т.д., и такие, которые сами по себе не определяют, какую – либо качественную реакцию или признак, а лишь усиливают или ослабляют проявление действия основного гена. Эти гены названы генами – модификаторами, а их действие – модифицирующим.
Однако существуют группы генов, которые отчетливо показывают своё модифицирующее действие на проявление отдельных генов. У таких генов – модификаторов часто не обнаруживается их самостоятельное действие на особь. Об их существовании мы узнаём по их влиянию на другие гены. По типу своего действия гены – модификаторы представлены двумя категориями: 1) гены, усиливающие проявление признака, детерминируемого другим геном; 2) гены, ослабляющие действие другого гена.
Особое значение приобретает действие генов – модификаторов в генетике количественных признаков.
Изменчивость по количественным признакам свойственна особям любой популяции внутри вида, сорта или породы. Отчасти эта изменчивость вызвана влиянием внешней среды, однако в большей мере её появление вызвано расщеплением по генам – модификаторам.
Гены, которые полностью или частично восстанавливают нормальное развитие особи, получили название супрессоры. Явление супрессии может быть результатом второй мутации в том же гене.
В присутствии супрессоров некоторые гены не могут проявлять своего действия. Такие супрессоры часто не оказывают никакого другого заметного влияния на фенотип. В широком смысле, по мнению Н.П. Дубинина (1986), в категорию супрессоров необходимо отнести и явление эпистаза.
7.3 Пенетрантность, экспрессивность
Рассматривая действие гена, его аллелей, необходимо учитывать не только генные взаимодействия, но и модифицирующее действие среды,в которой развивается организм.
Варьирующие соотношение классов при расщеплении в зависимости от условий внешней среды (так назвал С.С. Четвериков варьирование генотипа по генам – модификаторам) носит название варьирующей пенетрантности. Это понятие подразумевает возможность проявления или не проявления признака у организмов, одинаковых по исследуемым генотипическим факторам. Таким образом, пенетрантность выражается долей особей, проявляющих исследуемый признак среди всех особей одинакового генотипа по контролируемому (изучаемому) гену.
От внешней среды и генов – модификаторов может зависеть и степень выраженности признака. Например, дрозофила, гомозиготная по аллелям VgVg (зачаточные крылья), более контрастно проявляет этот признак при понижении температуры.
Степень проявления варьирующего признака называется экспрессивностью. Экспрессивность обычно выражается количественно в зависимости от уклонения данного типа.
Оба понятия – пенетрантность и экспрессивность – были введены для описания варьирующего проявления генов. Тот факт, что признак может проявляться или не проявляться у особей данного генотипа в зависимости от условий или варьировать в различных условиях среды, убеждает в том, что фенотип – это результат действия (и взаимодействия) генов в конкретных условиях существования организма.
ГЕНЕТИКА ПОЛА
8.1 Внеядерная цитоплазматическая наследственность
Хромосомная теория наследственности установила ведущую роль ядра и находящихся в них хромосом в явлениях наследственности. Но в то же время уже в первые годы формирования генетики как науки были известны факты, показывающие, что наследование некоторых признаков связано с нехромосомными компонентами клетки и не подчиняется менделевским закономерностям, основанным на распределении хромосом во время мейоза.
В 1908 – 1909 г.г. К. Корренс и одновременно независимо от него Э. Баур описали пёстролистность у растений ночной красавицы и львиного зева, которая наследуется через цитоплазму. В последующие годы подобные наблюдения были сделаны на других объектах. Все они правильно истолковывались, как примеры цитоплазматической наследственности.
Дальнейшее изучение явлений наследственности привело к необходимости установить не только механизм передачи генов, но и то, как эти гены контролируют процессы клеточного метаболизма и развитие определённых признаков и свойств. Поэтому клетку стали рассматривать, как единую целостную систему, определяющую передачу и воспроизведение признаков в потомстве в результате взаимодействия компонентов ядра (генов хромосом) и цитоплазмы, что можно показать на примере приобретения ею способности к фотосинтезу. Фотосинтез связан с цитоплазматическими структурами клетки – пластидами и находится в них пигментом – хлорофиллом.
К внеядерной наследственности относятся: цитоплазматическая и вирусная. Свидетельством цитоплазматической наследственности являются факты, ♀А х ♂В и обратном ♀В х ♂А направлениях. Потомство в обоих случаях похоже на материнские растения.
Образование и функции пластид обуславливаются наследственными факторами и действием внешних условий (главным образом света, без которого хлорофилл в пластидах не образуется). Таким образом, важнейшее свойство клетки – её способность к фотосинтезу – определяется взаимодействием генов хромосом, структурных элементов цитоплазмы и условий внешней среды. Схематический материал (по. Дж. Джинксу) можно представить так (рисунок 3).


 Геном Хромосомы Гены
Геном Хромосомы Гены
Поиск по сайту: