АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
Экологические пирамиды. Экосистемы очень разнообразны по относительной скорости создания и расходования, как первичной продукции
Экосистемы очень разнообразны по относительной скорости создания и расходования, как первичной продукции, так и вторичной продукции на каждом трофическом уровне. Однако, всем без исключения экосистемам свойственны определенные количественные соотношения первичной и вторичной продукции, получившие название правила пирамиды продукции: на каждом предыдущем трофическом уровне количество биомассы, создаваемой за единицу времени, больше, чем на последующем. Графически это правило выражают в виде пирамид, суживающихся кверху и образованных поставленными друг на друга прямоугольниками равной высоты, длина которых соответствует масштабам продукции на соответствующих трофических уровнях. Пирамида продукции отражает законы расходования энергии в пищевых цепях.[2]
Скорость создания органического вещества не определяет его суммарные запасы, т. е. общую биомассу всех организмов каждого трофического уровня. Наличная биомасса продуцентов или консументов в конкретных экосистемах зависит от того, как соотносятся между собой темпы накопления органического вещества на определенном трофическом уровне и передачи его на вышестоящий, т. е. насколько сильно выедание образовавшихся запасов. Немаловажную роль при этом играет скорость оборота генераций основных продуцентов и консументов.

Рис. 1. Пирамиды биомассы в некоторых биоценозах (по Ф. Дре, 1976): П – продуценты; РК – растительноядные консументы; ПК – плотоядные консументы; Ф – фитопланктон; 3 – зоопланктон
В большинстве наземных экосистем действует также правило пирамиды биомасс, т. е. суммарная масса растений оказывается больше, чем биомасса всех фитофагов и травоядных, а масса тех, в свою очередь, превышает массу всех хищников (рис. 1). Отношение годового прироста растительности к биомассе в наземных экосистемах сравнительно невелико. В разных фитоценозах, где основные продуценты различаются по длительности жизненного цикла, размерам и темпам роста, это соотношение варьирует от 2 до 76 %. Особенно низки темпы относительного прироста биомассы в лесах разных зон, где годовая продукция составляет лишь 2–6% от общей массы растений, накопленной в телах долгоживущих крупных деревьев. Даже в наиболее продуктивных дождевых тропических лесах эта величина не превышает 6,5 %. В сообществах с господством травянистых форм скорость воспроизводства биомассы гораздо выше: годовая продукция в степях составляет 41–55 %, а в травяных тугаях и эфемерно-кустарниковых полупустынях достигает даже 70–76 %. [2]
Отношение первичной продукции к биомассе растений определяет те масштабы выедания растительной массы, которые возможны в сообществе без подрыва его продуктивности. Относительная доля потребляемой животными первичной продукции в травянистых сообществах выше, чем в лесах. Копытные, грызуны, насекомые-фитофаги в степях используют до 70 % годового прироста растений, тогда как в лесах в среднем не более 10 %. Однако возможные пределы отчуждения растительной массы животными в наземных сообществах не реализуются полностью и значительная часть ежегодной продукции поступает в опад.
В пелагиали океанов, где основными продуцентами являются одноклеточные водоросли с высокой скоростью оборота генераций, их годовая продукция в десятки, и даже сотни, раз может превышать запас биомассы (рис. 2). Вся чистая первичная продукция так быстро вовлекается в цепи питания, что накопление биомассы водорослей очень мало, но вследствие высоких темпов размножения небольшой их запас оказывается достаточным для поддержания скорости воссоздания органического вещества.
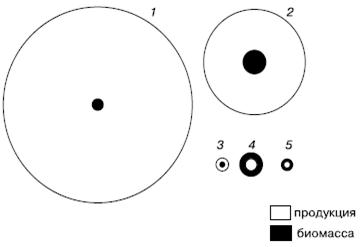
Рис. 2. Схема соотношения продукции и биомассы у бактерий (1), фитопланктона (2), зоопланктона (3), бентоса (4) и рыб (5) в Баренцевом море (по Л. А. Зенкевичу из С. А. Зернова, 1949)
Для океана правило пирамиды биомасс недействительно (пирамида имеет перевернутый вид). На высших трофических уровнях преобладает тенденция к накоплению биомассы, так как длительность жизни крупных хищников велика, скорость оборота их генераций, наоборот, мала и в их телах задерживается значительная часть вещества, поступающего по цепям питания.
В тех трофических цепях, где передача энергии происходит в основном через связи хищник – жертва, часто выдерживается правило пирамиды чисел: общее число особей, участвующих в цепях питания, с каждым звеном уменьшается. Это связано с тем, что хищники, как правило, крупнее объектов своего питания и для поддержания биомассы одного хищника нужно несколько или много жертв. Из этого правила могут быть и исключения – те редкие случаи, когда более мелкие хищники живут за счет групповой охоты на крупных животных. Правило пирамиды чисел было подмечено еще в 1927 г. Ч. Элтоном, который отметил также, что оно неприменимо к цепям питания паразитов, размеры которых с каждым звеном уменьшаются, а число особей возрастает.[2]
Пирамиды чисел позволяют не только сравнивать различные биоценозы, но и выявлять относительную значимость популяций в пределах одного сообщества. Они являются наиболее полезными из трех типов экологических пирамид, однако получить данные для их построения труднее всего.[3]
Все три правила пирамид – продукции, биомассы и чисел – выражают в конечном счете энергетические отношения в экосистемах, и если два последних проявляются в сообществах с определенной трофической структурой, то первое (пирамида продукции) имеет универсальный характер.[2]
Сообщества организмов связаны с неорганической средой теснейшими материально-энергетическими связями. Растения могут существовать только за счет постоянного поступления в них углекислого газа, воды, кислорода, минеральных солей. Гетеротрофы живут за счет автотрофов, но нуждаются в поступлении таких неорганических соединений, как кислород и вода. В любом конкретном местообитании запасов неорганических соединений, необходимых для поддержания жизнедеятельности населяющих его организмов, хватило бы ненадолго, если бы эти запасы не возобновлялись. Возврат биогенных элементов в среду происходит как в течение жизни организмов (в результате дыхания, экскреции, дефекации), так и после их смерти, в результате разложения трупов и растительных остатков. Таким образом, сообщество образует с неорганической средой определенную систему, в которой поток атомов, вызываемый жизнедеятельностью организмов, имеет тенденцию замыкаться в круговорот.
Знание законов продуктивности экосистем, возможность количественного учета потока энергии имеют чрезвычайное практическое значение. Первичная продукция агроценозов и эксплуатации человеком природных сообществ – основной источник запасов пищи для человечества. Не менее важна и вторичная продукция, получаемая за счет сельскохозяйственных и промысловых животных, так как животные белки включают целый ряд незаменимых для людей аминокислот, которых нет в растительной пище. Точные расчеты потока энергии и масштабов продуктивности экосистем позволяют регулировать в них круговорот веществ таким образом, чтобы добиваться наибольшего выхода выгодной для человека продукции. Кроме того, необходимо хорошо представлять допустимые пределы изъятия растительной и животной биомассы из природных систем, чтобы не подорвать их продуктивность. этом случае продолжать умеренно использовать.
Поиск по сайту: