АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
Доказательства генетической роли нуклеиновых кислот
Дымшиц
Лекция 1.
Что такое молекулярная биология. Доказательства генетической роли нуклеиновых кислот.
Определение предмета молекулярная биология
Термин "молекулярная биология" принадлежит Фрэнсису Крику, которому надоело в ответ на вопрос о его профессии объявлять себя смесью кристаллографа, биохимика, биофизика и генетика.
После атомной бомбежки Хиросимы и Нагасаки в 1945г. началось бегство ученых из физики, а в 1947г. Нобелевский лауреат физик Эрвин Шредингер написал книгу "Что такое жизнь с точки зрения физика?", которая привлекла в биологию многих физиков и математиков.
Определение: Mолекулярная биология - это наука о механизмах хранения, воспроизведения, передачи и реализации генетической информации, о структуре и функциях нерегулярных биополимеров - нуклеиновых кислот и белков.
Начав с изучения биологических процессов на молекулярно-атомном уровне, молекулярная биология перешла к сложным надмолекулярным клеточным структурам, а в настоящее время успешно решает проблемы генетики, физиологии, эволюции и экологии.
Основные открытия
| 1944г. | Доказательство генетической роли ДНК. Освальд Эйвери, Колин Мак-Леод, Маклин Мак-Карти |
| 1953г. | Установление структуры ДНК. Джеймс Уотсон, Френсис Крик |
| 1961г. | Открытие генетической регуляции синтеза ферментов. Андре Львов, Франсуа Жакоб, Жак Моно |
| 1962г. | Расшифровка генетического кода. Маршалл Нирнберг, Генрих Маттеи, Северо Очоа |
| 1967г. | Синтез in vitro биологически активной ДНК. Артур Корнберг (неформальный лидер молекулярной биологии) |
| 1970г. | Химический синтез гена. Гобинд Корана |
| 1970г. | Открытие фермента обратной транскриптазы и явления обратной транскрипции. Говард Темин, Дэвид Балтимор, Ренато Дульбеко |
| 1974г. | Открытие рестриктаз. Гамильтон Смит, Даниэль Натанс, Вернер Арбер |
| 1978г. | Открытие сплайсинга. Филипп Шарп |
| 1982г. | Открытие автосплайсинга. Томас Чек |
Доказательства генетической роли нуклеиновых кислот
1. 1928г. Опыты Фредерика Гриффита.

| Гриффит работал с пневмококками - бактериями, вызывающими пневмонию. Он брал два штамма пневмококков: капсульный и бескапсульный. Капсульный - патогенный (вирулентный), при инфицировании таким штаммом мыши погибают, бескапсульный - непатогенный. При введении мышам смеси убитых нагреванием (и, следовательно, потерявших вирулентность) капсульных пневмококков и живых бескапсульных невирулентных бактерий, животные погибали в результате размножения капсульных вирулентных форм. Обнаруженное явление Гриффит интерпретировал как трансформацию. |
Определение: Трансформация - это приобретение одним организмом некоторых признаков другого организма за счет захвата части его генетической информации.
В 1944г. этот эксперимент был повторен Освальдом Эйвери, Колином Мак-Леодом и Маклином Мак-Карти в варианте смешивания бескапсульных пневмококков с взятыми от капсульных белками, полисахаридами или ДНК. В результате этого эксперимента была выявлена природа трансформирующего фактора.
Трансформирующим фактором оказалась ДНК.
2. 1952г. Эксперимент Альфреда Херши и Марты Чейз.
Фаги (бактериофаги) - это вирусы, размножающиеся в бактериях.
E. сoli - кишечная палочка (эубактерия).

| Суть опыта: фаги, у которых белковая оболочка была мечена радиоактивной серой (S35), а ДНК - радиоактивным фосфором (Р32), инкубировали с бактериями. Затем бактерии отмывали. В смывных водах не обнаруживали Р32, а в бактериях - S35 Следовательно, внутрь попала только ДНК. Через несколько минут из бактерии выходили десятки полноценных фагов, содержащих и белковую оболочку, и ДНК. |
Отсюда следовал однозначный вывод о том, что именно ДНК выполняет генетическую функцию - несет информацию как о создании новых копий ДНК, так и о синтезе фаговых белков.
3. 1957г. Опыты Френкеля - Конрата

| Френкель-Конрат работал с вирусом табачной мозаики (ВТМ). В этом вирусе содержится РНК, а не ДНК. Было известно, что разные штаммы вируса вызывают разную картину поражения листьев табака. После смены белковой оболочки "переодетые" вирусы вызывали картину поражения, характерную для того штамма, чья РНК была покрыта чужим белком. |
Следовательно, не только ДНК, но и РНК может служить носителем генетической информации.
На сегодняшний день существуют сотни тысяч доказательств генетической роли нуклеиновых кислот. Приведенные три являются классическими.
1. Романтический период 1935-1944 гг.
Макс Дельбрюк и Сальвадор Лурия занимались изучением репродукции фагов и вирусов, представляющих собой комплексы нуклеиновых кислот с белками
В 1940г. Джордж Бидл и Эдуард Татум сформулировали гипотезу - "Один ген - один фермент". Однако, что такое ген в физико-химическом плане тогда еще не знали.
2. Второй романтический период 1944-1953 гг.
Была доказана генетическая роль ДНК. В 1953 г. появилась модель двойной спирали ДНК, за которую ее создатели Джеймс Уотсон, Френсис Крик и Морис Уилкинс были удостоены Нобелевской премии.
3. Догматический период 1953-1962 гг. Сформулирована центральная догма молекулярной биологии:
Перенос генетической информации идет в направлении ДНК →РНК →белок
В 1962 г. был расшифрован генетический код.
4. Академический период с 1962 г. по настоящее время, в котором с 1974 года выделяют генно-инженерный подпериод.
Лекция 2. Строение нуклеиновых кислот
Хронология открытий, подготовивших создание Уотсоном и Криком модели двойной спирали ДНК
| 1868г. | Обнаружен нуклеин. Современное название - хроматин. Фридрих Мишер |
| 1889г. | Нуклеин разделен на нуклеиновую кислоту и белок. Появился термин "нуклеиновая кислота". Рихард Альтман |
| 1900г. | Все азотистые основания были описаны химиками. |
| 1909г. | В нуклеиновых кислотах обнаружены фосфорная кислота и рибоза. Левин |
| 1930г. | Найдена дезоксирибоза. Левин |
| 1938г. | Рентгеноструктурный анализ показал, что расстояние между нуклеотидами в ДНК 3,4 Å. При этом азотистые основания уложены стопками. Уильям Астбюри, Флорин Белл |
| 1947г. | С помощью прямого и обратного титрования установлено, что в ДНК есть водородные связи между группами N-H и C=O. Гулланд |
| 1953г. | С помощью кислотного гидролиза ДНК с последующей хроматографией и количественным анализом установлены закономерности: А/Т=1; Г/Ц=1; (Г+Ц)/(А+Т)=К - коэффициент специфичности, постоянен для каждого вида. Эрвин Чаргафф |
Правила Чаргаффа. В ДНК всегда А/Т=1; Г/Ц=1; (Г+Ц)/(А+Т)=К - коэффициент специфичности, постоянен для каждого вида.
Нуклеиновые кислоты являются нерегулярными полимерами, мономеры которых - нуклеотиды.

| Нуклеотид = нуклеозид + фосфорная кислота = азотистое основание+ пентоза+фосфорная кислота. В РНК пентоза - рибоза. В ДНК - дезоксирибоза. |

| Нуклеотиды соединяются друг с другом в полимерную цепочку с помощью фосфодиэфирных связей. Азотистые основания не принимают участия в соединении нуклеотидов одной цепи. |

| Существует два класса азотистых оснований. Пурины: аденин (А) и гуанин (Г) - содержат два гетероцикла. Пиримидины: тимин (Т), цитозин (Ц) и урацил (У) - содержат один гетероцикл. |
Принципы строения ДНК

|
1. Нерегулярность.
Существует регулярный сахарофосфатный остов, к которому присоединены азотистые основания. Их чередование нерегулярно.
2. Антипараллельность.
ДНК состоит из двух полинуклеотидных цепей, ориентированных антипараллельно. 3`-конец одной расположен напротив 5`-конца другой.
3. Комплементарность (дополнительность).
Каждому азотистому основанию одной цепи соответствует строго определенное азотистое основание другой цепи. Соответствие задается химией. Пурин и пиримидин в паре образуют водородные связи. В паре A-Т две водородные связи, в паре Г-Ц - три.
4. Наличие регулярной вторичной структуры.
Две комплементарные, антипараллельно расположенные полинуклеотидные цепи образуют правые спирали с общей осью.
Формы двойной спирали ДНК
Существуют несколько форм двойной спирали ДНК.

|
В основной - В-форме на виток приходится 10 комплементарных пар. Плоскости азотистых оснований перпендикулярны оси спирали. Соседние комплементарные пары повернуты друг относительно друга на 36  . Диаметр спирали 20Å, причем пуриновый нуклеотид занимает 12Å, а пиримидиновый - 8Å. А-форма - 11 пар азотистых оснований на виток. Плоскости азотистых оснований отклонены от нормали к оси спирали на 20 . Отсюда следует наличие внутренней пустоты диаметром 5Å. Высота витка 28Å. Такие же параметры у гибрида из одной цепи ДНК и одной цепи РНК.
. Диаметр спирали 20Å, причем пуриновый нуклеотид занимает 12Å, а пиримидиновый - 8Å. А-форма - 11 пар азотистых оснований на виток. Плоскости азотистых оснований отклонены от нормали к оси спирали на 20 . Отсюда следует наличие внутренней пустоты диаметром 5Å. Высота витка 28Å. Такие же параметры у гибрида из одной цепи ДНК и одной цепи РНК.
С-форма - шаг спирали 31Å, 9.3 пар оснований на виток, угол наклона к перпендикуляру 6 .
Все три формы - правозакрученные спирали.
Есть еще несколько форм правых спиралей и всего одна левая спираль (Z -форма). Высота витка в Z-форме -44.5 Å, на виток приходится 12 пар нуклеотидов. Ни А-, ни Z- формы не могут существовать в водном растворе без дополнительных воздействий (белки или суперспирализация).
Отличия между ДНК и РНК
| ДНК | РНК | |
| Сахар | Дезоксирибоза | Рибоза |
| Азотистые основания | А, Т, Г, Ц | А, У, Г, Ц |
| Количество цепей в молекуле | 99.99% двойная спираль 0.01% одноцепочечная. | 99.99% одноцепочечная 0.01% двухцепочечная |
| Форма молекулы | Все одноцепочечные- кольцевые. Большинство двухцепочечных - линейные, часть- кольцевые. | Линейные молекулы |
Виды РНК
| Виды РНК | Размер в нуклеотидах |
| gРНК - геномные РНК | 10000-100000 |
| mРНК - информационные (матричные) РНК | 100-100000 |
| tPHK - транспортные РНК | 70-90 |
| rРНК - рибосомные РНК | несколько дискретных классов от 100 до 500000 |
| sРНК - малые РНК | 100-300 |
Функции ДНК
1. ДНК является носителем генетической информации.
Функция обеспечивается фактом существования генетического кода.
2. Воспроизведение и передача генетической информации в поколениях клеток и организмов.
Функция обеспечивается процессом репликации.
3. Реализация генетической информации в виде белков, а также любых других соединений, образующихся с помощью белков-ферментов.
Функция обеспечивается процессами транскрипции и трансляции.
Лекция 3. Генетический код
Определение: Генетический код - это система записи информации о последовательности расположения аминокислот в белках с помощью последовательности расположения нуклеотидов в ДНК.
Поскольку ДНК непосредственного участия в синтезе белка не принимает, то код записывается на языке РНК. В РНК вместо тимина входит урацил.
Свойства генетического кода
1. Триплетность
Каждая аминокислота кодируется последовательностью из 3-х нуклеотидов.
Определение: триплет или кодон - последовательность из трех нуклеотидов, кодирующая одну аминокислоту.

Код не может быть моноплетным, поскольку 4 (число разных нуклеотидов в ДНК) меньше 20. Код не может быть дуплетным, т.к. 16 (число сочетаний и перестановок из 4-х нуклеотидов по 2) меньше 20. Код может быть триплетным, т.к. 64 (число сочетаний и перестановок из 4-х по 3) больше 20.
2. Вырожденность.
Все аминокислоты, за исключением метионина и триптофана, кодируются более чем одним триплетом:
2 АК по 1 триплету = 2
9 АК по 2 триплета = 18
1 АК 3 триплета = 3
5 АК по 4 триплета = 20
3 АК по 6 триплетов = 18
Всего 61 триплет кодирует 20 аминокислот.
3. Наличие межгенных знаков препинания.
Определение: ген- это участок ДНК, кодирующий одну полипептидную цепь или одну молекулу tРНК, rРНК или sРНК.
Гены tРНК, rРНК, sРНК белки не кодируют.
В конце каждого гена, кодирующего полипептид, находится, по меньшей мере, один из 3-х терминирующих кодонов, или стоп-сигналов: UAA, UAG, UGA. Они терминируют трансляцию.
Условно к знакам препинания относится и кодон AUG - первый после лидерной последовательности. (См. лекцию 8) Он выполняет функцию заглавной буквы. В этой позиции он кодирует формилметионин (у прокариот).
4. Однозначность.
Каждый триплет кодирует лишь одну аминокислоту или является терминатором трансляции.
Исключение составляет кодон AUG. У прокариот в первой позиции (заглавная буква) он кодирует формилметионин, а в любой другой - метионин.
5. Компактность, или отсутствие внутригенных знаков препинания.
Внутри гена каждый нуклеотид входит в состав значащего кодона.
В 1961г. Сеймур Бензер и Френсис Крик экспериментально доказали триплетность кода и его компактость.

| Суть эксперимента: "+" мутация - вставка одного нуклеотида. "-" мутация - выпадение одного нуклеотида. Одиночная "+" или "-" мутация в начале гена портит весь ген. Двойная "+" или "-" мутация тоже портит весь ген. Тройная "+" или "-" мутация в начале гена портит лишь его часть. Четверная "+" или "-" мутация опять портит весь ген. Эксперимент доказывает, что код триплетен и внутри гена нет знаков препинания. Эксперимент был проведен на двух рядом расположенных фаговых генах и показал, кроме того, наличие знаков препинания между генами. |
6. Универсальность.
Генетический код един для всех живущих на Земле существ.
Это является сильнейшим свидетельством в пользу единства происхождения и эволюции.
В 1979г. Беррел открыл идеальный код митохондрий человека.
Определение: идеальным называется генетический код, в котором выполняется правило вырожденности квазидублетного кода:
Если в двух триплетах совпадают первые два нуклеотида, а третьи нуклеотиды относятся к одному классу (оба - пурины или оба - пиримидины), то эти триплеты кодируют одну и ту же аминокислоту.
Из этого правила в универсальном коде есть два исключения. Оба отклонения от идеального кода в универсальном касаются принципиальных моментов: начала и конца синтеза белка:
| Кодон | Универсальный код | Митохондриальные коды | |||
| Позвоночные | Беспозвоночные | Дрожжи | Растения | ||
| UGA | STOP | Trp | Trp | Trp | STOP |
| AUA | Ile | Met | Met | Met | Ile |
| CUA | Leu | Leu | Leu | Thr | Leu |
| AGA | Arg | STOP | Ser | Arg | Arg |
| AGG | Arg | STOP | Ser | Arg | Arg |
7. Помехоустойчивость.
Определение: Мутации замен нуклеотидов, не приводящие к смене класса кодируемой аминокислоты, называют консервативными. Мутации замен нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют радикальными.
В каждом триплете можно провести 9 однократных замен. Общее количество возможных замен нуклеотидов - 61 по 9 = 549. Из них:
23 замены нуклеотидов приводят к появлению кодонов - терминаторов трансляции.
134 замены не меняют кодируемую аминокислоту.
230 замен не меняют класс кодируемой аминокислоты.
162 замены приводят к смене класса аминокислоты, т.е. являются радикальными.
Из 183 замен 3-его нуклеотида, 7 приводят к появлению терминаторов трансляции, а 176 - консервативны.
Из 183 замен 1-ого нуклеотида, 9 приводят к появлению терминаторов, 114 - консервативны и 60 - радикальны.
Из 183 замен 2-го нуклеотида, 7 приводят к появлению терминаторов, 74 - консервативны, 102 - радикальны.
Итак: 364/162=2.25 (отношение числа консервативных замен к числу радикальных замен) - показатель помехоустойчивости генетического кода.
8. Неперекрываемость.

| В 1956г. Георгий Гамов предложил вариант перекрываемого кода. Согласно Гамовскому коду, каждый нуклеотид, начиная с третьего в гене, входит в состав 3-х кодонов. Когда генетический код был расшифрован, оказалось, что он неперекрываем, т.е. каждый нуклеотид входит в состав лишь одного кодона. |
Достоинства перекрываемого генетического кода:
компактность, меньшая зависимость структуры белка от вставки или делеции нуклеотида.
Недостаток:
большая зависимость структуры белка от замены нуклеотида и ограничение на соседей.

| В 1976г. была секвенирована ДНК фага φХ174. У него одноцепочечная кольцевая ДНК, состоящая из 5375 нуклеотидов. Было известно, что фаг кодирует 9 белков. Для 6 из них были определены гены, располагающиеся друг за другом. Выяснилось, что есть перекрывание. Ген Е полностью находится внутри гена D. Его инициирующий кодон появляется в результате сдвига считывания на один нуклеотид. Ген J начинается там, где кончается ген D. Инициирующий кодон гена J перекрывается с терминирующим кодоном гена D в результате сдвига на два нуклеотида. Конструкция называется "сдвиг рамки считывания" на число нуклеотидов, некратное трем. На сегодняшний день перекрывание показано только для нескольких фагов. |
Информационная емкость ДНК
На Земле живет 6 миллиардов человек. Наследственная информация о них заключена в 6х109 сперматозоидах. По разным оценкам у человека от 30 до 50 тысяч генов. У всех людей ~ 30х1013 генов или 30х1016 пар нуклеотидов, которые составляют 1017 кодонов. Средняя книжная страница содержит 25х102 знаков. ДНК 6х109 сперматозоидов содержит информацию, равную по объему примерно 4х1013 книжных страниц. Эти страницы заняли бы объем 6-и зданий НГУ. 6х109 сперматозоидов занимают половину наперстка. Их ДНК занимает менее четверти наперстка.
Лекция 4. Транскрипция
Определение: транскрипция - это синтез всех видов РНК по матрице ДНК, осуществляемый ферментом ДНК-зависимой РНК-полимеразой.
У прокариот синтез всех видов РНК осуществляется одним и тем же ферментом.
У эукариот - 3 ядерные РНК-полимеразы, митохондриальные РНК-полимеразы, хлоропластные РНК-полимеразы.
Субстратами для РНК-полимераз служат рибонуклеозид-трифосфаты (активированные нуклеотиды). Весь процесс транскрипции осуществляется за счет энергии макроэргических связей актвированных нуклеотидов.

Принципы транскрипции:
1. Комплементарность.
2. А нтипараллельность.
3. Униполярность.
4. Беззатравочность.
5. Асимметричность.
РНК синтезируется комплементарно и антипараллельно транскрибируемой цепи ДНК. Рост цепи РНК идет только в направлении 5'  3'. Для начала синтеза РНК фермент не нуждается в поли- или олигонуклеотидной затравке.
3'. Для начала синтеза РНК фермент не нуждается в поли- или олигонуклеотидной затравке.
Первый нуклеотид в РНК всегда пурин в форме трифосфата.
Асимметричность
Транскрибируются обе цепи ДНК, но в каждом отдельном опероне только одна из них. Какая именно, определяется положением промотора и терминатора.
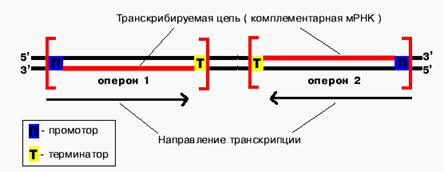
Понятие об опероне
Определение: оперон - единица транскрипции у прокариот.
В начале каждого оперона находится промотор.В конце каждого оперона находится терминатор. Перед терминатором располагаются структурные гены, или цистроны. Между промотором и цистронами может находиться оператор. Существуют моно-, олиго- и полицистронные опероны.

Определение: промотор - особая последовательность нуклеотидов ДНК, узнаваемая РНК-полимеразой как посадочная площадка и старт синтеза РНК.
Только с промотора может начаться синтез специфической РНК.
Определение: терминатор - особая последовательность нуклеотидов ДНК, узнаваемая РНК-полимеразой как финиш транскрипции.
Определение: цистрон - последовательность нуклеотидов ДНК, кодирующая один полипептид (в большинстве случаев - белок) или одну tРНК, или одну rРНК.
В большинстве случаев цистроны объединяются в оперон по следующему принципу: закодированные в них белки принимают участие в одной биохимической цепи реакций.
Определение: оператор - особая последовательность нуклеотидов ДНК, узнаваемая белком-репрессором.
У оператора диспетчерская функция - он разрешает или запрещает синтез РНК.

Субъединичный состав РНК-полимеразы Е.coli
РНК-полимераза Е.coli - белок с четвертичной структурой. Одновременно в клетке присутствует около 7000 молекул РНК-полимеразы.
Субъединичный состав фермента: (2  )
) 
 - holo -фермент (полный фермент). Без -фактора это core -фермент (2 ) .
- holo -фермент (полный фермент). Без -фактора это core -фермент (2 ) .
(сигма) - фактор - сменный фактор специфичности.
Только holo -фермент обладает высоким сродством к специфической последовательности нуклеотидов - промотору, сродство к остальным случайным последовательностям ДНК у него снижено в 10000 раз. У core -фермента одинаковое сродство к любой последовательности нуклеотидов.
Сам по себе - фактор обладает наименьшим сродством к ДНК по сравнению с другими субьединицами РНК-полимеразы, однако он придает holo -ферменту такую конформацию, которая обладает повышенным сродством к промотору.
Как только произошла инициация транскрипции, -фактор отделяется. Элонгация - продолжение синтеза РНК, и терминация - его остановка, осуществляются core -ферментом.
Стадии узнавания и связывания, а также инициации осуществляются holo -ферментом. Элонгация и терминация осуществляются core -ферментом.
Две субъединицы - каркас РНК-полимеразы. К ним крепятся остальные субъединицы.
- субъединица отвечает за прочное связывание с ДНК за счет кластера положительно заряженных аминокислот.
В - субъединице находятся два каталитических центра. Один отвечает за инициацию, а другой - за элонгацию. Один центр работает в holo -, а другой - в core - ферменте.
Особенности структуры промотора
Для изучения структуры промоторов провели следующий эксперимент. При оптимальных условиях связывания получили комплекс РНК-полимеразы с ДНК. Этот комплекс обработали ДНК-азой, и таким образом гидролизовали всю ДНК, незащищенную РНК-полимеразой. После этого отделили РНК-полимеразу от оставшихся фрагментов ДНК. Опять создали оптимальные условия для образования комплекса. Комплекс не образовывался.

Отсюда следует вывод, что
узнавание и прочное связывание происходит на разных участках ДНК.
Эти участки отличаются и по первичной, и по вторичной структуре. Путем секвенирования выявили структуру многих промоторов. У большинства из них имеется общее свойство.

РНК-полимераза узнает промотор, покрывая 40-60 пар нуклеотидов. В промоторе узнается взаимное расположение двух расплавленных AT-богатых участков. В каждом из них расплавлено 4-6 пар. Центры этих участков находятся в положенях "-10" и "-35". Принципиально важным является расстояние между расплавленными участками. Оно колеблется от 16 до 19 п.н. Искусственное увеличение этого расстояния до 20 п.н. или уменьшение его до 15 п.н. приводит к тому, что РНК-полимераза не узнает испорченный промотор.
Этапы транскрипции
1. Узнавание и прочное связывание
Как только произошло узнавание (позиция 1), РНК-полимераза перемещается в позицию 2. В каталитическом центре инициации транскрипции, находящемся в -субъединице, оказывается +1-ый нуклеотид оперона. Переход из позиции 1 в позицию 2 возможен, если на операторе нет белка-репрессора.

Примерно 5% промоторов у прокариот имеют только участок "-10", однако, тем не менее, хорошо узнаются РНК-полимеразой. Такие промоторы представлены палиндромными последовательностями, принимающими форму креста при суперспирализации кольцевых молекул ДНК.
Определение: палиндромы - последовательности, которые читаются одинаково слева направо и справа налево.
Палиндромы первого порядка имеют одну ось симметрии, второго - две, третьего - три.

2. Инициация заключается в образовании первой фосфодиэфирной связи между пурин-трифосфатом (АТФ или ГТФ) и следующим нуклеотидом. После инициации - фактор покидает фермент.
3. Элонгация - последовательное наращивание цепи РНК (или продолжение транскрипции).
Скорость элонгации 40-50 нукл./сек.
Для комплементарного синтеза РНК необходим разрыв водородных связей в ДНК. Core -фермент РНК- полимеразы покрывает примерно 40 пар нуклеотидов (4 витка спирали ДНК). Разрыв водородных связей на 4-х витках спирали - очень энергоемкий процесс. Он не был обнаружен при изучении транскрипции.

| Показано, что РНК-полимераза переводит ДНК из В-формы в А-форму. В ней плоскости азотистых оснований не перпендикулярны оси спирали, а наклонены на 200 к перпендикуляру. Это облегчает "выворачивание" двух соседних азотистых оснований в цепи ДНК для того, чтобы напротив них встали комплементарные нуклеотиды РНК. В пользу этого говорит полная идентичность параметров А-формы ДНК и гибрида, состоящего из одной цепи ДНК и одной - РНК. "Мотором" транскрипции является энергия, высвобождающаяся при отщеплении пирофосфата от каждого рибо-НТФ. |
4. Терминация.
Специфическая терминация бывает  - независимой и - зависимой.
- независимой и - зависимой.

| При - независимой терминации в терминаторе присутствует палиндром. В синтезируемой РНК формируется шпилька. Шпилька меняет конформацию РНК-полимеразы и фермент теряет сродство к ДНК.
|
- зависимая терминация.
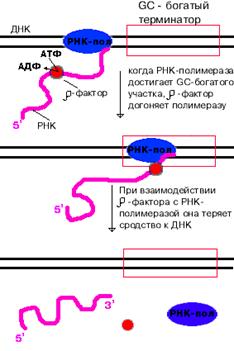
| - фактор - это имеющий четвертичную структуру белок, обладающий АТФ-азной активностью. Он способен узнавать 5`-конец синтезируемой РНК длиной приблизительно 50 нуклеотидов, садиться на него и двигаться по РНК с такой же скоростью, с которой РНК-полимераза движется по ДНК.
В терминаторе много Г-Ц пар (с тремя водородными связями), вследствие чего РНК-полимераза замедляет ход, - фактор ее догоняет, изменяет конформацию фермента - и синтез РНК прекращаетс
|
Ингибиторы транскрипции прокариот.
Существует множество ингибиторов транскрипции. Они действуют по разным механизмам и на разных стадиях. Большинство из них - антибиотики.
Рифампицин - ингибитор инициации.
Связывается с центром инициации holo -РНК-полимеразы E. сoli.
Стрептолидигин - ингибитор элонгации.
Связывается с центром элонгации core -РНК-полимеразы E. сoli.
Лекция 5. Регуляция транскрипции у прокариот
Позитивный контроль работы lac -оперона
Lac -оперон, подчиняющийся схеме негативной индукции, имеет и позитивный контроль.
| цАМФ образуется из АТФ ферментом аденилатциклазой. Фосфодиэстераза превращает цАМФ в АМФ. Глюкоза активирует второй и инактивирует первый фермент. Чем больше в клетке глюкозы, тем меньше цАМФ. | 
| |
| Если нет глюкозы, то цАМФ соединяется с белком катаболической репрессии (САР) и образуется комплекс САР·цАМФ, активирующий посадку РНК-полимеразы на промотор. В присутствии лактозы lac -оперон включается и работает. | 
| |
Если же в клетке есть еще и глюкоза (более экономичный источнок энергии), то нет цАМФ - и активатор не образуется, lac -оперон работает "вяло", без дополнительной индукции.
Схема негативной индукции Жакоба и Моно
Lac -оперон E. coli содержит 3 гена, отвечающие за образование белков, участвующих в переносе в клетку дисахарида лактозы и в ее расщеплении.
Z - - галактозидаза (расщепляет лактозу на глюкозу и галактозу).
Y- - галактозидпермеаза (переносит лактозу через мембрану клетки).
А - тиогалактозидтрансацетилаза (ацетилирует галактозу).
| В отсутствие в клетке лактозы lac - оперон выключен. Активный белок - репрессор, кодируемый в моноцистронном опероне (LacI), не имеющем оператора, связан с оператором lac -оперона. Поскольку оператор перекрывается с промотором, даже посадка РНК-полимеразы на промотор невозможна. | 
|
| Как только некоторое количество лактозы попадает в клетку, две молекулы субстрата (лактозы) взаимодействуют с белком - репрессором, изменяют его конформацию - и он теряеет сродство к оператору. Тут же начинается транскрипция lac -оперона и трансляция образующейся mРНК; три синтезируемых белка участвуют в утилизации лактозы. |
Когда вся лактоза переработана, очередная порция репрессора, свободного от лактозы, выключает lac -оперон.
Эта схема называется так потому, что контролирующим транскрипцию фактором является негативный фактор, "выключатель" - белок - репрессор. Индукция (включение) происходит при потере сродства белка - репрессора к оператору.
Существует и позитивная регуляция работы lac -оперона E. coli.
Схема негативной репрессии
Оперон синтеза триптофана у E. сoli.
| В опероне имеется 5 цистронов, которые кодируют ферменты последовательной цепи реакций синтеза триптофана. В норме оперон включен. Белок - репрессор неактивен (в форме апо-репрессора), он не способен садиться на оператор. | 
|
| Клетке нужно N молекул триптофана. N+1-ая молекула взаимодействует с апо-репрессором. Он меняет конформацию, садится на оператор и синтез РНК прекращается. | 
|
Схема регуляции - негативная репрессия, потому что белок репрессор "выключает" оперон.
Помимо "грубой схемы" включения - выключения, есть и тонкая регуляция синтеза триптофана - аттенуация(см. лекцию N 9).
Схема позитивной индукции
Аra -оперон E. сoli.
| В нем 3 цистрона, которые кодируют ферменты, расщепляющие сахар арабинозу. В норме оперон закрыт. Белок - репрессор связан с оператором. | 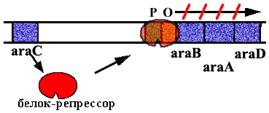
|
| Когда в клетку попадает арабиноза, она взаимодействует с белком - репрессором. Белок - репрессор меняет конформацию и превращается из репрессора в активатор, взаимодейсивующий с промотором и облегчающий посадку РНК-полимеразы на промотор. | 
|
Эта схема регуляции называется позитивной индукцией, поскольку контролирующий элемент - белок - активатор "включает" работу оперона.
Схема позитивной репрессии
Оперон синтеза рибофлавина у Вacilus subtilis.
| В опероне располагаются цистроны ферментов синтеза рибофлавина. Есть белок-активатор, обеспечивающий посадку РНК-полимеразы на промотор. В норме оперон открыт. Образуется N молекул рибофлавина. | 
|
| N+1-ая молекула (лишняя) взаимодействует с активатором и он теряет способность активировать посадку РНК-полимеразы на промотор. | 
|
Позитивная репрессия, поскольку в регуляции участвует белок - активатор, а сама регуляция заключается в выключении транскрипции.
Лекция 6. Транскрипция у эукариот
У эукариот процессы транскрипции и трансляции разобщены во времени и пространстве (транскрипция - в ядре, трансляция - в цитоплазме).
У эукариот существуют специализированные РНК-полимеразы.
В ядре выделяют 3 типа РНК-полимераз:
РНК-полимераза I - синтезирует rРНК (кроме 5S rРНК).
РНК-полимераза II - синтезирует mРНК и некоторые sРНК.
РНК-полимераза III - синтезирует tРНК, некоторые sРНК и 5SrРНК.
РНК-полимеразы различаются количеством субъединиц, их аминокислотным составом, и зависимостью от катионов магния и марганца. Для РНК-полимераз I и III необходимое для работы соотношение [Mn2+]/[Mg2+] = 2. Для РНК-полимеразы II - [Mn2+]/[Mg2+] = 5.
Наиболее яркое различие - чувствительность к - аманитину (токсину бледной поганки). Он полностью подавляет работу РНК-полимеразы II в концентрации 10-8 М и РНК-полимеразы III (в концентрации 10-6 М). РНК-полимераза I фактически нечувствительна к этому токсину.
Помимо ядерных РНК-полимераз у эукариот есть еще РНК-полимеразы хлоропластов и митохондрий. Они кодируются в ядре, а не в соответствующих органеллах.
В органеллах образуются свои tРНК, rРНК и рибосомные белки.
Как образуются рибосомы у эукариот
Гены rРНК присутствуют в количестве от 10 до 105 копий у разных видов (105 у амфибий). У человека - 300 генов, в которых закодированы rРНК.
Все рибосомные гены, кроме генов 5S рибосомной РНК, сближены (т.е располагаются один за другим) и образуют несколько кластеров. Сначала синтезируется про-rРНК, после созревания которой образуются 28S, 18S и 5,8S rРНК.

|
Интерфазные хромосомы в световой микроскоп не видны. Каждый ген прорибосомной РНК транскрибируется одновременно несколькими РНК-полимеразами и тут же начинается процессинг.
На электронномикроскопических фотографиях видна картина "рождественской елочки". Синтезируемые в ядре mРНК поступают на готовые рибосомы в цитоплазму, где синтезируются рибосомные белки, которые идут в ядро и путаются в "ветвях елки".
Образуются рибосомные субъединицы. Одновременно в эукариотическом ядре находятся сотни тысяч субъединиц рибосом.
Определение: ядрышко - место образования субъединиц рибосом, наблюдаемое в световой микроскоп.
В ядре может быть несколько ядрышек.
Определение: кластер генов rРНК называют ядрышковым организатором.
Особенности транскрипции эукариот
Единицей транскрипции у эукариот является отдельный ген, а не оперон, как у прокариот.
Оператор, как таковой, отсутствует.
Промотор есть, но он организован иначе.
На расстоянии -25 п.н. от +1 нукл. находится ТАТА-бокс. Его позиция определяет точку инициации транскрипции. А на расстоянии -60-80 п.н. находится ЦААТ-бокс, который не является абсолютно необходимым, но присутствует перед большинством генов.
Расстояние между ЦААТ и ТАТА большое и РНК-полимераза не способна накрыть всю эту область.
ЦААТ опознается своим белком, а ТАТА - своим.

|
Помимо этих есть еще несколько белков, называемых базальными факторами транскрипции.
Определение: базальные факторы транскрипции - белки, необходимые для инициации транскрипции.
Базальные факторы транскрипции необходимы для инициации транскрипции всеми тремя ядерными РНК-полимеразами.
Для любого гена, кодирующего белок, есть энхансеры (усилители).
Определение: энхансеры - последовательности ДНК, усиливающие транскрипцию при взаимодействии со специфическими белками.
| Энхансеры - это не непрерывные последовательности нуклеотидов. Существуют так называемые модули - это отдельные части энхансеров. Одинаковые модули могут встречаться в разных энхансерах. Для каждого энхансера набор модулей уникален. Модули - это короткие последовательности, не более 2-х витков спирали (20 п.н.), которые могут находиться перед, за и даже внутри гена. | 
|
Таким образом, М1+М2+М3+М4 - один энхансер, но он состоит из 4-х модулей. Все 4 модуля узнаются своими белками, а они, сидя на ДНК, взаимодействуют друг с другом. Если в клетке присутствуют все соответствующие белки, то участку ДНК придается определенная конформация и начинается синтез mРНК.
Все соматические клетки многоклеточного эукариотического организма имеют абсолютно одинаковый набор генов. Почему же клетки дифференцированы и специализированы?
Дело в том, что все гены работают на фоновом уровне и не имеют фенотипического проявления. Экспрессируются лишь те гены, у которых все энхансерные модули узнаны своими белками и эти белки взаимодействуют друг с другом. Кроме энхансеров есть сайленсеры (ослабители).
Определение: сайленсеры - последовательности ДНК, ослабляющие транскрипцию при взаимодействии с белками.
При соответствующем наборе белков экспрессия отдельных генов в клетке может быть подавлена.
Процессинг mРНК
Процессинг (созревание) rРНК и tРНК у эукариот принципиально не отличается от такового у прокариот.
Процессинг mРНК отличается сильно и состоит из нескольких этапов.
1. Кепирование 100% mРНК
2. Полиаденилирование ~95% mРНК
3. Сплайсинг ~95% mРНК. Сплайсингу подвергаются только полиаденилированные mРНК.
4. Редактирование
Показано лишь для нескольких mРНК.
Все стадии процессинга mРНК происходят в РНП-частицах (рибонуклеопротеидных комплексах.
По мере синтеза про-mРНК, она тут же образует комплексы с ядерными белками - информоферами. И в ядерные, и в цитоплазматические комплексы mРНК с белками (информосомы) входят sРНК.
Таким образом, mРНК не бывает свободной от белков.
На всем пути следования до завершения трансляции mРНК защищена от нуклеаз. Кроме того, белки придают ей необходимую конформацию.
Определение: полисома - комплекс mРНК с несколькими или многими рибосомами.
В составе информосом mРНК может жить от нескольких минут до нескольких дней, не подвергаясь действию нуклеаз. (Так, mРНК живут неделями в ооцитах, предшественниках яйцеклеток).
Кепирование
Кепирование - надевание "шапочки".

|
"Сар" представляет собой метилированный ГТФ, присоединенный в необычной позиции 5'-5' и две метилированные рибозы в первых двух нуклеотидах mРНК. По мере образования про-mРНК (еще до 30-ого нуклеотида), к 5'-концу, несущему пуринтрифосфат, присоединяется гуанин, после чего происходит метилирование.
ГТФ + гуанинтрансфераза (Е) Е~фГ + ф-ф
5'ф-ф-ф-Пур-ф-Х-ф-Y-ф-Z-...+E~фГ Г5'-ф-ф-ф-5'-Пур-ф-Х-ф-Y-ф-Z...
(метилирование) Гm-ф-ф-ф-Пурm-ф-Хm-ф-Y-ф-Z-...
Назначение "Сар"
1. Защита 5'-конца mРНК от действия экзонуклеаз.
2. За счет узнавания "Сар" -связывающими белками происходит п равильная установка mРНК на рибосоме.
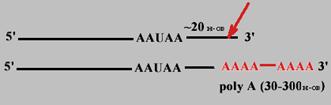
Полиаденилирование
|
Когда синтез про-mРНК завершен, то на расстоянии примерно 20 нуклеотидов в направлении к 3' - концу от последовательности 5'-AAУAA-3' происходит разрезание специфической эндонуклеазой и к новому 3'-концу присоединяется от 30 до 300 остатков АМФ (безматричный синтез).
Каждый вид mРНК имеет "поли-А хвост" определенной длины. Он защищает 3'-конец от гидролиза, т.к. покрыт полиА-связывающими белками.
В значительной степени время жизни mРНК коррелирует с длиной полиА-хвоста.
mРНК ряда генов не полиаденилируется (например гистоновых генов).
Полиаденилированные про-mРНК подвергаются сплайсингу.
Сплайсинг
В 1978г. Филипп Шарп (Массачусетский технологический институт) открыл явление сплайсинга РНК (от англ. to splace - сшивать без узлов).
Определение: экзоны - кодирующие участки генов.
Определение: интроны - некодирующие участки генов.
На долю интронов приходится в 5-7 раз больше нуклеотидных пар, чем на долю экзонов. Количество экзонов в гене больше, чем интронов.

|
Определение: сплайсинг - вырезание копий интронов из про-mРНК и сшивание копий экзонов с образованием mРНК.
Копии интронов гидролизуются до нуклеотидов.
Сплайсинг показан для большинства mРНК и некоторых tРНК. У простейших найден автосплайсинг rРНК. Сплайсинг показан даже для археобактерий.
Не существует единого механизма сплайсинга. Описано по крайней мере 5 разных механизмов.
В ряде случаев сплайсинг осуществляют ферменты-матюразы.
В некоторых случаях в процессе сплайсинга участвуют sРНК.
В случае автосплайсинга процесс происходит благодаря третичной структуре про-РНК.
Для mРНК высших организмов существуют обязательные правила сплайсинга:
Правило 1. 5' и 3' концы интрона очень консервативны: 5'(ГT-интрон-AГ)3'.

|
Правило 2. При сшивании копий экзонов соблюдается порядок их расположения в гене, но могут быть выброшены некоторые из них.

|
Представление об интроне, как пустой, ничего не кодирующей последовательности, неверно. Некоторые интроны кодируют ферменты-матюразы, вырезающие копии этих интронов. На вопрос, зачем эукариотическим геномам экзон - интронная структура, можно ответить только в единичных случаях. Почему эукариотические гены "разорваны", будет рассмотрено в конце курса.
Альтернативный сплайсинг mРНК кальцитонинового гена у млекопитающих (крыса)
Во всех клетках есть кальцитониновый ген, но в клетках щитовидной железы он экспрессируется в виде гормона кальцитонина, а в клетках гипофиза - нейропептида CGRP (пептида, имеющего отношение к гену кальцитонина). Ген один, а белки получаются разные в результате сплайсинга mРНК и процессинга полипептидов. В клетках других тканей этот ген не экспрессируется.
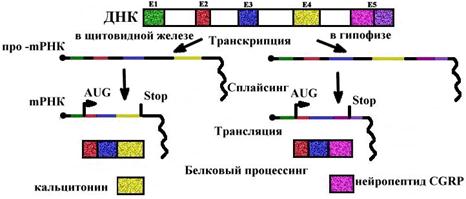
|
Сплайсинг осуществляется белковыми комплексами - сплайсосомами, в которых помимо ферментов, вырезающих и сшивающих участки про-mРНК, имеются белки, придающие про-mРНК нужную конформацию, и несколько sPНК. Сплайсосома непосредственно связана с ферментами, занимающимися полиаденилированием.
Автосплайсинг
Автосплайсинг открыт Томасом Чеком (США) в 1982 году.
Он работал с инфузорией Tetrаchymenа thermophyla. У этой инфузории образуется 35S про-rРНК длиной 6400 нуклеотидов. Без участия дополнительных соединений белковой природы из этой про-rРНК вырезается внутренний участок длиной в 414 нуклеотида. Два экзона сшиваются с образованием 26S rРНК. Единственное требование - определенная концентрация ионов магния. Про-rРНК имеет третичную структуру и обладает каталитичекой активностью. Впервые было показано, что каталитической активностью обладают не только белки.

|
Определение: РНК-зимы - РНК с каталитической активностью.
Сегодня описано несколько десятков РНК-зимов.
Редактирование
Определение: редактирование - изменение генетической информации на уровне mРНК.
Трипаносома - одноклеточный паразит, вызывающий у человека сонную болезнь. В клетке трипаносомы есть митохондрии и множество ферментов окислительного фосфорилирования. Один из ключевых ферментов - цитохромоксидаза. Он имеет четвертичную структуру и состоит из 3-х разных субъединиц.
Человек (постоянный хозяин трипаносомы) теплокровен, поэтому трипаносоме не нужна энергия собственных митохондрий. Синтезируются только две субъединицы цитохромоксидазы.
Муха цеце (промежуточный хозяин трипаносомы) - холоднокровна. Трипаносоме нужен работающий фермент. Все три субъединицы синтезируются.
В геноме трипаносомы только два гена для двух субъединиц. В mРНК одного из них происходит разрезание и встраивание 4-х урацилов (на небольшом расстоянии друг от друга, но не подряд).
Происходит сдвиг рамки считывания и отредактированная mРНК кодирует новый полипептид - третью субъединицу цитохромоксидазы.

|
Малые РНК
sРНК обнаружены в количестве 103-105 копий на клетку. Поскольку в большинстве случаев эти РНК обогащены урацилом, они называются U1, U2... Их размер от 100 до 300 нукл. Все они кодируются в ядре, но работают как в ядре (small nuclear - SN), так и в цитоплазме (small cytoplasmic - SC).
SNURPS - РНП (рибонуклеопротеидные комплексы) в ядре. snРНК входят в состав РНП, участвующих в полиаденилированиии и сплайсинге.
SCURPS - РНП в цитоплазме. Входят в состав информосом.
Малая РНК U4 присутствует в комплексах, участвующих в полиаденилировании. Если получить антитела к белкам, связывающимся с U4, то не происходит полиаденилирования и сплайсинга.
При красной волчанке (аутоимунном заболевании) вырабатываются антитела к белкам комплекса с U4.
Гистоновая mРНК не полиаденилируется потому, что sРНКU7, которая комплементарна 3'-концу гистоновой mРНК, защищает ее от полиаденилирования.
Малые РНК U1, U2, U4, U5, U6 входят в состав сплайсосомы.
| Лекция 7. Репликация. |
Определение: процесс, осуществляемый комплексом ферментов и белков, выполняющих топологическую функцию, суть которого в образовании идентичных копий ДНК для передачи генетической информации в поколениях клеток и организмов, называют репликацией ДНК.
Принципы репликации
1. Комплементарность.
2. Антипараллельность.
3. Униполярность.
4. Потребность в затравке.
5. Прерывистость.
6. Полуконсервативность.
Первые три принципа можно сформулировать в одной фразе:
Синтез каждой дочерней цепи ДНК идет комплементарно и антипараллельно матричной цепи и всегда в направлении 5' 3'.
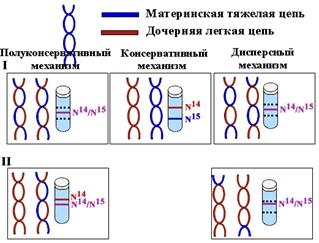
Доказательство полуконсервативного характера репликации
Для выяснения вопроса о характере расхождения цепей по дочерним молекулам Мэтт Мезельсон и Фрэнк Сталь в 1958г. разработали метод равновесного центрифугирования в градиенте плотности CsCl..
ДНК разделяется не по молекулярным весам, а по удельной плотности.
E. сoli выращивали на протяжении нескольких поколений на среде, содержащей тяжелый изотоп азота (N15), для того, чтобы вся ДНК была "тяжелой". Перед очередным раундом деления клетки синхронизировали. При этом в среде заменяли N15 на легкий изотоп N14 с тем, чтобы вновь синтезированные цепи были "легкими". После репликации ДНК выделяли и центрифугировали в градиенте плотности CsCl.
Полуконсервативность означает, что каждая дочерняя ДНК состоит из одной матричной цепи и одной вновь синтезированной.
|
Равное распределение "тяжелых" и "легких" цепей между всеми молекулами исключало возможность консервативного способа, согласно которому одна дочерняя клетка получает материнскую ДНК, а другая - вновь синтезированную, обе цепи которой являются новыми.
Клетки второго поколения содержали как полностью "легкие" молекулы, так и "гибридные", состоящие из одной "легкой" и одной "тяжелой" цепи, аналогичные молекулам первого поколения. Этот факт исключал возможность дисперсного механизма, согластно которому куски материнской ДНК случайным образом распределяются между дочерними молекулами.
Ферментативная система синтеза ДНК in vitro
В 1956 г. Артур Корнберг наработал 100 кг биомассы E. coli и выделил 0.5 г фермента ДНК-полимеразы.
Необходимые компоненты для синтеза ДНК in vitro:
1. ДНК-матрица - образец, по которому строится новая цепь ДНК.
2. Активированные нуклеотиды (dАТФ, dГТФ, dТТФ, dЦТФ) - то, из чего строятся дочерние цепи.
3. ДНК-полимераза - то, что строит новую цепь ДНК.
4. Ионы магния - то, без чего фермент не работает.
ДНК-матрицу необходимо активировать.
Нативная двуцепочечная ДНК, не имеющая повреждений, не может эффективно использоваться в этой системе. Активировать ее можно либо денатурацией щелочью или нагреванием (1), либо обработкой экзонуклеазой III из E. сoli (2), либо внесением ников (одноцепочечных разрывов) с помощью эндонуклеаз (3).

|
Строение и свойства ДНК-полимеразы Корнберга (ДНК-полимеразы I)
ДНК - полимераза Корнберга (ДНК-полимераза I) - это одна полипептидная цепь с молекулярным весом 109 тыс.
В состав полимеразы входят ионы цинка. Она абсолютно зависима от ионов магния.
Обнаружено 4 разные каталитические активности ДНК - полимеразы I:
1. Полимеризационная в направлении 5` 3`.
ДНКn + dХТФ ДНКn+1 + Ф-Ф
2. Пирофосфоролиз:
ДНКn + Ф-Ф* ДНКn-1 + dХТФ*
Эта активность возможна при большом избытке пирофосфатов.
3. Пирофосфатный обмен:
ДНКn + dХТФ + Ф-Ф* ДНКn+ dХТФ* + Ф-Ф
Фермент работает только тогда, когда он находится на молекуле ДНК и имеет соответствующую конформацию.
4. Гидролитическая активность:
ДНКn + Н2О ДНКn-1 + dХMФ
Эта активность имеет большой биологический смысл.
Гидролитическая активность проявляется в направлении 3' 5' и 5' 3'.
Активность 3' 5' проявляется на неспаренном 3'-гидроксильном конце. Фермент возвращается при ошибке включения и "откусывает" неправильный нуклеотид.
Это корректорская функция фермента.
Все ДНК-полимеразы обладают этой активностью.
Фермент способен гидролизовать спаренный 5'-конец, расчищая себе дорогу и продолжая полимеризацию.

| Если на пути фермента встречается короткий (меньше 10 нуклеотидов) неспаренный 5'-конец, то полимераза сначала проявляет эндонуклеазную активность и откусывает весь свисающий конец, а затем проявляет экзонуклеазную 5' 3' активность т.е. откусывает по одному нуклеотиду.
Если неспаренный 5'-конец длинный, то фермент использует его как матрицу.
| |
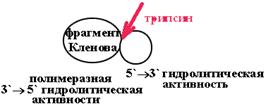
| При мягком расщеплении ДНК-полимеразы трипсином можно получить два активных фрагмента: один обладает полимеразной и 3' 5' гидролитической активностью (фрагмент Кленова), другой - 5' 3' гидролитической активностью.
| 
| |
Понятие о матрице и затравке
Продукты, образуемые в ферментативной системе in vitro.
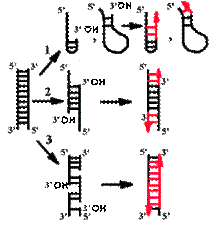
|
Во всех случаях матрицей для синтеза новых цепей служит одноцепочечная ДНК. Затравкой является 3'-гидроксильный конец двуцепочечной ДНК, причем он должен быть спарен с матрицей.
В том случае, если эндонуклеаза вносила ники с 3'-фосфатным концом, ДНК не являлась активированной.
Прямым доказательством того, что затравка - 3'-гидроксильный конец, является эксперимент с дидезоксинуклеозидтрифосфатом.

| Если такой активированный нуклеотид сделать меченым по α-фосфату, то он включается в растущую полимерную цепь и всегда обнаруживается на ее 3'-конце. Это говорит о том, что он сам включается, но дальнейший рост цепи невозможен, т.к. нет 3'-гидроксильного конца. |
Это также доказывает и униполярность репликации в направлении 5' 3'.
Схема размножения фага М13
 1974 г. Оказаки.
1974 г. Оказаки.
Рифампицин - ингибитор бактериальной РНК-полимеразы (на стадии инициации).
Хлорамфеникол - ингибитор трансляции на бактериальных рибосомах.
Если одновременно с заражением E. сoli фагом добавить хлорамфеникол, то блокируются трансляция, репликация II и сборка фагов.
Если подействовать рифампицином, то блокируется не только транскрипция и все следующие процессы, но и репликация I.
Вывод: бактериальная РНК-полимераза участвует в репликации ДНК фага.
Вся фаговая ДНК составляет ~6000 нукл.
Определение: origin (ori) - район начала репликации.

| В районе ori (начало репликации) имеется 4 шпильки. Эти шпильки опознаются РНК-полимеразой, и вторая шпилька используется в качестве матрицы. По мере образования РНК шпилька плавится. Образуется РНК-затравка длиной 24 нукл., 3'-конец которой используется ДНК полимеразой III. |
| Когда 3'-конец синтезируемой цепи ДНК "утыкается" в 5'-конец РНК-затравки, ДНК-полимераза III вытесняется ДНК-полимеразой I, которая, обладая 5' 3' гидролитической активностью, "съедает" РНК-затравку, одновременно продолжая синтез ДНК. Когда затравка (РНК) съедена, ДНК-полимераза I вытесняется лигазой, которая сшивает концы ДНК.
| 
|
Все эти ферменты (ДНК-полимераза III, ДНК-полимераза I, лигаза) входят в состав реплисомы. Они представляют единый белковый комплекс, который реагирует изменением конформации на выполнение очередной функции.
Протяженная (более 100 нукл.) одноцепочечная ДНК-матрица может быть использована ДНК полимеразой III, когда полимераза III представлена в форме holo -фермента. Помимо субъединицы , обладающей полимеразной активностью, в holo -фермент входит еще несколько субъединиц, обеспечивающих высокую процессивность синтеза ДНК.
Фаг 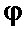 X174
X174
Репликация ДНК этого фага не зависит от рифампицина.
Поиск по сайту: