АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
Влияние тяжелых металлов на микроорганизмы в почве
Тяжелые металлы существенным образом влияют на численность, видовой состав и жизнедеятельность почвенной микробиоты. Они ингибируют процессы минерализации и синтеза различных веществ в почвах, подавляют дыхание почвенных микроорганизмов, вызывают микробостатический эффект и могут выступать как мутагенный фактор.
Большинство тяжелых металлов в повышенных концентрациях ингибируют активность ферментов в почвах: амилазы, дегидрогеназы, уреазы, инвертазы, каталазы. На основании этого предложены индексы, аналогичные широко известному показателю ЛД50, в которых действующей считается концентрация загрязнителя, на 50 или на 25% снижающая определенную физиологическую активность, например уменьшение выделения СО2 почвой – ЭкД50, ингибирование активности дегидрогеназы - EC50, подавление активности инвертазы на 25 %, снижение активности восстановления трехвалентного железа – ЕД50.
Токсичность тяжелых металлов неодинаково проявляется по отношению к различным ферментам. В дерново-подзолистой почве наиболее чувствительной оказалась каталаза, активность которой снижалась на 25% при концентрации кадмия, в 3 раза превышающей фоновую цинка - в 10 раз, свинца - в 25 раз, Практически нечувствительной к изученным металлам была уреаза. Некоторые авторы наиболее чувствительными называют разные ферменты: инвертазу, дегидрогеназу, уреазу.
По степени токсичности в отношении ферментов тяжелые металлы формируют следующий убывающий ряд: Cd > Pb > Zn.
При изучении влияния тяжелых металлов в полевых условиях закономерности обнаружить значительно сложнее, вследствие высокой вариабельности естественной природной среды. В результате во многих работах не обнаружено достоверных изменений нитрифицирующей и азотфиксирующей активностей, выделения углекислого газа, активности ферментов. С другой стороны, отмечается, что активность азотфиксации может в значительной степени подавляться ТМ, другие процессы цикла азота слабее ингибируются этими элементами. Показано снижение активности азотфиксации в почвах под хвойными лесами при действии меди, никеля и цинка, в дерново-подзолистой, торфяно-глеевой почвах и черноземе типичном при действии кадмия и ртути. Свинец оказывал менее заметный эффект, а в малых концентрациях даже стимулировал азотфиксацию.
Цинк, свинец, кадмий, медь и ртуть также подавляли азотфиксацию серозема обыкновенного, наиболее четко это проявлялось при концентрациях металлов, в 10-50 и более раз превышающих фоновые. С увеличением концентраций металлов закономерно снижалась пространственная вариабельность данного процесса.
Тяжелые металлы могут вызывать изменения общей численности почвенных микроорганизмов. Во многих работах показано снижение количества прокариотных микроорганизмов в разных типах почв: подзолистой, торфянистой, дерново-подзолистой, черноземе выщелоченном, сероземе обыкновенном, черноземе типичном и других. Часто отмечается снижение численности олигонитрофильных и аммонифицирующих бактерий, коринеподобных и некоторых споровых бактерий, актиномицетов. Относительно более высокая устойчивость характерна для целлюлозолитических бактерий. Количество микроскопических грибов в загрязненных тяжелыми металлами почвах, наоборот, может возрастать.
Наиболее чувствительной группой по результатам лабораторных опытов в черноземе типичном оказались липомицеты, плотность популяции которых значительно снижалась уже при 5-ти кратном превышении фоновых концентраций тяжелых металлов. В торфяно-глеевой почве чувствительным организмом оказался Azotobacter chroococum, численность которого резко снижалась при 20-ти кратном превышении фоновой концентрации загрязнителей. В дерново-подзолистой почве ни одна из изученных групп микроорганизмов не проявила сравнительно высоких индикационных свойств. В целом, 50%-ное снижение численности всех трех изученных групп микроорганизмов достигается лишь при 100-ти кратном превышении фонового содержания тяжелых металлов в каждом из трех изученных типов почв.
В полевых условиях данные по численности сложнее использовать из-за большой природной вариабельности этого показателя. В связи с этим вполне логичными выглядят сообщения ряда авторов об отсутствии достоверных изменений количества микроорганизмов в загрязненных почвах. Такие данные приводятся для микробиоты лесной почвы, количества грамотрицательных бактерий и актиномицетов в почве под соей, плотности популяций бактерий, грибов и актиномицетов, численности азотобактера, грамположительных бактерий. Некоторые авторы полагают, что по численности микроорганизмов (споровых бактерий, грибов, актиномицетов) невозможно обнаружить существенных отличий в действии тяжелых металлов на почвы разных типов в концентрациях, в несколько раз превышающих фоновые.
Загрязнение почв тяжелыми металлами вызывает определенные изменения в видовом составе комплекса почвенных микроорганизмов. В качестве общей закономерности отмечается значительное сокращение видового богатства и разнообразия комплекса почвенных микромицетов при загрязнении. Кроме того, в грибном сообществе загрязненной почвы появляются необычные для нормальных условий, по-видимому, устойчивые к тяжелым металлам виды микромицетов.
Различную чувствительность к тяжелым металлам проявляют виды почвенных простейших и водорослей.
Показано, что наиболее чувствительными к загрязнению являются виды рода Вacilllls, несколько более устойчивы к металлам псевдомонады, стрептомицеты и многие виды целлюлозоразрушающих микроорганизмов.
Грамотрицательные бактерии, особенно Flavobacterium spp., Serratia spp. и некоторые другие представители этой группы более толерантны к металлам, чем грамположительные. На основании таких результатов предлагается использовать видовой состав почвенных микроорганизмов в качестве критерия для качественной оценки загрязняющего действия тяжелых металлов в почве.
Четкие изменения комплекса почвенных микромицетов под влиянием загрязнения ртутью проявляются при концентрациях, равных 50 мг/кг почвы, что более чем в 300 раз превышает уровень фона. Из 9 видов грибов, присутствовавших в контрольной почве, остаются только два вида (Trichoderma viride и Мuсог plumbeus) и обнаруживаются с высокой частотой встречаемости 3 вида (Penicillium skrjabinii, Aspergillus ustus и А. niger).
В полевых условиях на примере серозема обыкновенного, загрязненного тяжелыми металлами (в концентрациях, превышающих фон в 50-200 раз), также обнаружено заметное по частоте встречаемости видов изменение состава различных групп почвенных микроорганизмов, Так, комплекс почвенных микромицетов в незагрязненном сероземе включал 15 видов, частота встречаемости которых была более или менее равномерно распределена между всеми видами. В загрязненной почве богатство видов сокращается до 9, причем в их число входят 3 новых по сравнению с незагрязненным сероземом вида. Абсолютным доминантам по встречаемости в комплексе микромицетов становится вид Penicillium purpurogenum, встречаемость которого превышает 93%.
Таким образом, при сильном загрязнении почвы происходит снижение видового разнообразия комплекса микроорганизмов и увеличение абсолютного доминирования небольшого числа видов.
Изучение видового состава почвенных микроорганизмов позволило обнаружить ряд факторов, которые помогают понять некоторые негативные аспекты загрязняющего действия тяжелых металлов. Так, в некоторых случаях из загрязненных почв с высокой частотой встречаемости выделяют виды микромицетов, обладающие фитотоксическими свойствами (в частности, Penicillium purpurogenum).
Видовой состав дрожжей в почве также существенным образом меняется при загрязнении. Эти изменения выражаются в резком (на 40%) снижении доли типичных почвенных дрожжей в загрязненном сероземе и возрастании на 30% численности эпифитных дрожжей в названной почве. В результате количество почвенных дрожжей в незагрязненном сероземе более чем в 10 раз превышает число эпифитных, в то время как в загрязненной почве их пропорция составляет примерно 1,4. Увеличение количества эпифитных дрожжей, по нашему мнению, свидетельствует об угнетенном состоянии растительности на загрязненных территориях, что также может рассматриваться как негативный результат загрязнения.
Видовой состав дрожжей в почве также существенным образом меняется при загрязнении. Эти изменения выражаются в резком (на 40%) снижении доли типичных почвенных дрожжей в загрязненном сероземе и возрастании на 30% численности эпифитных дрожжей в названной почве. В результате количество почвенных дрожжей в незагрязненном сероземе более чем в 10 раз превышает число эпифитных, в то время как в загрязненной почве их пропорция составляет примерно 1,4. Увеличение количества эпифитных дрожжей, по нашему мнению, свидетельствует об угнетенном состоянии растительности на загрязненных территориях, что также может рассматриваться как негативный результат загрязнения.
В качестве общей закономерности отмечается, что повышенной толерантностью к различным экстремальным условиям окружающей среды, в том числе к тяжелым металлам, обладают пигментированные микроорганизмы или те, которые выделяют пигменты в среду. Предполагается, что эти соединения могут связывать металлы в нетоксичные комплексы. Особенно это относится к пигментам меланоидной природы.
При изучении секционного состава почвенных актиномицетов рода Streptomyces в загрязненном сероземе, содержащем тяжелые металлы в концентрации, в 50-200 раз превышающей фоновую, обнаружено изменение в соотношении отдельных секций. Снижается доля неокрашенных и слабоокрашенных актиномицетов секций Albus и Flavus. Доля окрашенных стрептомицетов (Chromogenes, Rubro-aurantiacus), наоборот, возрастает при загрязнении. Подобные результаты получены и при изучении комплекса почвенных микромицетов в загрязненном сероземе, где возрастает частота встречаемости видов с яркой пигментацией Penicillium purpurogenum и Р. herquei, меланизированного Aureobasidium pullulans, и дрожжей, где увеличивалась доля окрашенных видов Cryptococcus laurentii, Rhodotorula spp., Rhodosporidium spp. и некоторых других.
Таким образом, исследование изменений видового состава некоторых групп почвенных микроорганизмов под влиянием загрязнения показало, что этот фактор вызывает изменения в комплексе почвенных микроорганизмов. Это выражается в снижении видового богатства и разнообразия, увеличении доли толерантных к загрязнению микроорганизмов.
Исследование реакции на различные факторы окружающей среды активно функционирующих в почве микробных сообществ интенсивно развивается. Данный подход включает методы инициированного микробного сообщества, мембранных камер, двухслойных нестерильных почвенных пластинок и ряд других методов. Сходство этих методов заключается в том, что в почву вносится определенный питательный субстрат, который, как предполагается, стимулирует развитие наиболее активных группировок почвенных микроорганизмов. В дальнейшем изучается действие на эти микроорганизмы антропогенных нагрузок в градиенте концентраций, чтобы выявить закономерности типа доза-эффект.
Амилолитическое микробное сообщество дерново-подзолистой почвы устойчиво к внесению свинца в интервале концентраций до 200 мг на 1 кг воздушно-сухой почвы (рис. 18). Организация (степень доминирования отдельных членов сообщества) и состав (список видов сообщества) до этой дозы практически не изменялись.
Биомасса сообщества в пределах данного интервала концентраций загрязнителя несколько возрастала. При концентрациях свинца более 200 мг/кг в организации микробного сообщества происходили значительные изменения. Заметно снижалась доля доминирующих организмов сообщества контрольной почвы - Slreptomyces sp. 1 cfr. griseus, Coniothyrium fuckeli, Chaetomium globosum. Возрастала доля до этого редко встречавшихся микроорганизмов - Streptomyces sp. 2 cfr. Griseus, Monilia geophila, Arthrobotrys oligospora, Trichurus spirales, Bacillus polymyxa.
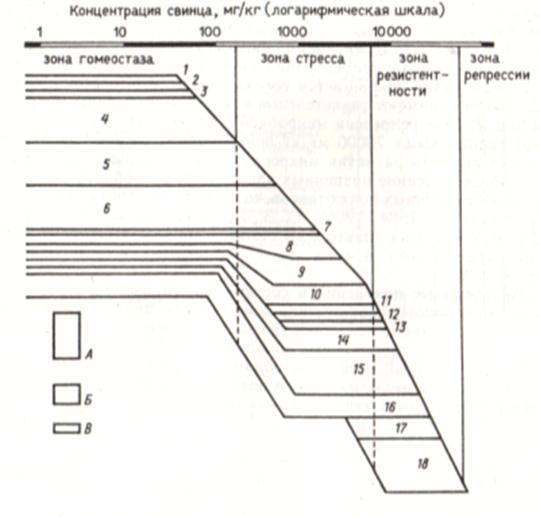
Рис. 18. Влияние свинца (Pb(CH3COO)2) на организацию амилолитического микробного сообщества дерново-подзолистой почвы:
А – доминирующие виды; Б – часто встречающиеся виды; В – редко встречающиеся виды.
1 – Streptomyces sp. 1 cfr. albus; 2 - Streptomyces sp. 2 cfr. albus; 3 – Nocardia sp.; 4 - Streptomyces sp. 1 cfr. griseus; 5 - Coniothyrium fuckeli; 6 - Chaetomium globosum; 7 – Mucor plumbeus; 8 - Streptomyces sp. 2 cfr. griseus; 9 - Monilia geophila; 10 – Arthrobotrys oligospora; 11 - Streptomyces sp. cfr. violaceus; 12 - Streptomyces sp. cfr. viride; 13 - Streptomyces sp. cfr. flavus; 14 – Trichurus spirales; 15 - Bacillus polymyxa; 16 – Stachybotrys altenans; 17 – Aspergillus flavus; 18 – Penecillium notatum
В составе сообщества как редко встречающиеся появлялись некоторые виды микроорганизмов, ранее не обнаруженные в контрольной почве. Среди них микроскопические грибы - Aspergillus flavus и Penicillium notatum. Эти организмы начинали активно развиваться при концентрации свинца около 7000 мг/кг. Общая биомасса микроорганизмов в интервале концентраций от 200 до 7000 мг Рb/кг проявляла значительную вариабельность.
После 10000 мг свинца доминировал Р. notatum, а часто встречающимся являлся вид А. flavus. Биомасса микроорганизмов сообщества в интервале концентраций свинца от 7000 до 10000 мг/кг почвы могла быть довольно значительной, но затем постепенно снижалась и около 70 тыс. мг Рb/кг практически равна нулю.
При последующем увеличении концентраций свинца, вносимых в почву, не удавалось обнаружить развития микроорганизмов.
В общем виде зависимость между градиентом концентраций свинца и изменениями микробного сообщества напоминает реакцию чистых культур микроорганизмов на возрастающие концентрации тяжелых металлов. При низких концентрациях тяжелых металлов наблюдается некоторая стимуляция развития инициированного сообщества микроорганизмов, затем частичное ингибирование и, наконец, полное подавление. Существенное отличие представленных выше результатов от данных, полученных в опытах с чистыми культурами, заключается в изменениях состава и организации микробного сообщества в градиенте концентраций тяжелого металла. Микроорганизмы, входящие в состав сообщества, обладают разной степенью устойчивости к тяжелым металлам и находятся в постоянном взаимодействии с почвой и между собой. Поэтому суммарный результат реакции всего инициированного микробного сообщества на тяжелые металлы определяется взаимодействием этих элементов с почвой, их действием на микроорганизмы и на конкурентные взаимоотношения микроорганизмов между собой. Доминирование разных микроорганизмов при различных концентрациях свинца позволило разделить весь диапазон толерантности сообщества на несколько участков - адаптивных зон, характеризующихся качественно отличными показателями (организацией и составом сообщества). Под адаптивной зоной понимается интервал концентраций изучаемого агента, определяющий совокупность изменений активно функционирующего в почве микробного сообщества, которые обеспечивают его приспособление к определенным условиям существования.
Адаптивная зона гомеостаза микробной системы почвы охватывает диапазон концентраций свинца от 0 до 200 мг/кг почвы, в котором изменяется только интенсивность микробиологических· процессов, что выражается, например, в возрастании суммарной биомассы сообщества. Зона стресса - диапазон концентраций от 200 до 7000 мг Рb/кг, где происходят существенные изменения организации инициированного микробного сообщества, вызванные перераспределением его членов по степени доминирования. Зона резистентности охватывает интервал концентраций свинца от 7000 до 70000 мг /кг, В котором резко сокращается состав сообщества, а доминирующее положение занимают резистентные к загрязнителю виды микроорганизмов. В зоне репрессии микробной системы почв (при концентрациях свинца выше 70000 мг/кг почвы) прямыми наблюдениями не удается выявить развития микроорганизмов на поверхности почвы.
Аналогичные изменения в составе и организации амилолитического микробного сообщества, описанные в виде вышеуказанных четырех адаптивных зон; обнаружены и при внесении других тяжелых металлов в некоторые почвы зонального ряда: сильноподзолистую, чернозем типичный и серозем обыкновенный. Те же четыре зоны были обнаружены и при внесении в дерново-подзолистую почву таких элементов, как кадмий, медь, никель и ртуть, в виде различных соединений: солей разной степени растворимости, оксидов. Таким образом, представленные результаты позволяют считать четырехэтапную реакцию инициированного микробного сообщества на загрязнение почвы антропогенными агентами достаточно универсальным явлением.
Аналогичные изменения в составе и организации амилолитического микробного сообщества, описанные в виде вышеуказанных четырех адаптивных зон; обнаружены и при внесении других тяжелых металлов в некоторые почвы зонального ряда: сильноподзолистую, чернозем типичный и серозем обыкновенный. Те же четыре зоны были обнаружены и при внесении в дерново-подзолистую почву таких элементов, как кадмий, медь, никель и ртуть, в виде различных соединений: солей разной степени растворимости, оксидов. Таким образом, представленные результаты позволяют считать четырехэтапную реакцию инициированного микробного сообщества на загрязнение почвы антропогенными агентами достаточно универсальным явлением.
Наиболее важной характеристикой, количественно отражающей степень устойчивости почвенной микробной системы к тяжелым металлам, является величина зоны гомеостаза. Говоря о значении величины зоны гомеостаза, следует иметь в виду верхнюю границу интервала концентраций загрязнителей, соответствующего этой зоне. При сравнении различных почв между собой по величине зон гомеостаза видно, что они выстраиваются в зонально-генетический ряд с максимумом устойчивости в черноземе и минимумом в подзолистой почве (рис. 19). Зона гомеостаза в черноземе в 10 раз шире, чем в подзолистой почве, что соответствует зональным особенностям этих почв: количеству гумуса, поглотительной способности почв, их буферной емкости и почвенному плодородию.
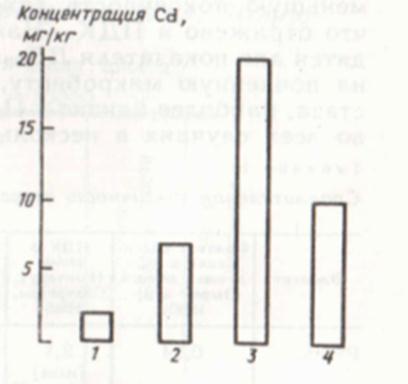
Рис. 19. Величина зоны гомеостаза микробной системы сильноподзолистой почвы (1), дерново-подзолистой почвы (2) чернозема типичного (3) и серозема обыкновенного (4) при загрязнении их кадмием (Cd(CH3COO)2)
Сказанное позволяет рассматривать величину зоны гомеостаза как объективный критерий, по которому можно сравнивать действие тяжелых металлов на микроорганизмы в различных почвах и количественно оценивать потенциальную устойчивость микробной системы к загрязнению. Этот же показатель может быть использован для изучения сравнительной физиологической активности различных элементов и форм их соединений в конкретной почве.
Полученные результаты по измерению величины зоны гомеостаза в дерново-подзолистой почве позволяют расположить соединения тяжелых металлов таким образом, что они выстраиваются в ряд с убывающей активностью. В общем виде этот ряд выглядит следующим образом: Hg>Cd>Ni>Cu>Pb.
Разные соединения ТМ влияют на почвенную микробиоту неодинаково. Как правило, их токсичность зависит от растворимости. Подвижные формы этих элементов обладают большей активностью, чем слаборастворимые.
Методом инициированного микробного сообщества изучали серозем обыкновенный, загрязненный тяжелыми металлами в условиях промышленного производства. Эта почва содержала концентрации тяжелых металлов, в 50-100 раз превышающие фоновое содержание.
В качестве контроля использовали близкую по всем характеристикам почву того же типа, но содержащую только фоновые концентрации тяжелых металлов. При внесении свинца в серозем с фоновым содержанием тяжелых металлов амилолитическое микробное сообщество сохраняет стабильность до 200 мг Рb/кг почвы (рис. 20).

Рис. 20. Влияние свинца (Pb(CH3COO)2) на организацию амилолитического микробного сообщества серозема обыкновенного (контрольная почва):
А – доминирующие виды; Б – часто встречающиеся виды; В – редко встречающиеся виды.
1 – Streptomyces sp. 1 cfr. flavus; 2 - Streptomyces sp. 1 cfr. griseus; 3 – Trichurus gorgonifer; 4 – Acremonium charticola; 5 - Chaetomium globosum; 6 - Monilia geophila; 7 - Aspergillus ustus; 8 - Streptomyces sp. cfr. albus; 9 - Coniothyrium macrocladium; 10 - Stachybotrys altenans; 11 - Penecillium funiculosum; 12 - Streptomyces sp. cfr. rubro-aurantiacus; 13 – Chaetomium anahelicinum; 14 – Monocillium indicum; 15 – Stysanus stemonites; 16 - Aspergillus flavus; 17 – Sepedonium chrysospermum
При увеличении концентраций свинца выше этой дозы снижается доля доминирующих организмов исходного сообщества - Streptomyces sp. cfr. flavus и Chaetomium globosum, возрастает доля ранее редко встречавшихся микроорганизмов: Aspergillus ustus, Streptomyces sp. cfr. albus, Streplomyces sp. cfr. rubro-aurantiacus, Chaelomium anahelicinum, Monocillium indicum, которые становятся часто встречающимися или доминирующими. Начиная с 20000 мг Рb/кг состав сообщества резко видоизменяется по сравнению с исходным. В него входят всего три вида, не обнаруживаемые в начальном сообществе и, по-видимому, толерантные к повышенным концентрациям свинца в почве: Slysanus stemonites в качестве редко встречающегося и Aspergillus flavus, Sepedonium chrysospermum - доминанты.
С 70000 мг Рb/кг ни на крахмальной полоске, ни на почве практически не удается обнаружить роста и развития микроорганизмов.
Таким образом, в контрольной почве обнаружены 4 адаптивные зоны микробной системы почвы.
Амилолитическое микробное сообщество серозема обыкновенного, загрязненного тяжелыми металлами в условиях промышленного производства (рис. 21), еще до лабораторного внесения свинца содержало в качестве доминанта актиномицет Streptomyces sp. cfr. rubrо-aurantiacus, часто встречающимися микроорганизмами были: Stysanus stemonites, Monocillium indicum, Streptomyces sp. cfr. albus, а среди редко встречающихся организмов был, например, Streptomyces sp. cfг. flavus. Такая организация инициированного микробного сообщества характерна для зоны стресса сообщества контрольной почвы (см. рис. 20). Состав сообщества сравнительно мало изменяется до внесения 3000 мг Рb/кг. При последующем увеличении доз свинца состав сообщества значительно изменяется: он становится беднее видами, но при этом появляются некоторые виды, не обнаруживаемые в сообществе контрольной почвы. Это, например, Streptoverlicillium sp., Penicillium notatum, доминирующие позиции занимают популяции микроорганизмов - Aspergillus flavus и Sepedonium chrysospermum. Начиная с 70000 мг Рb/кг почвы, роста микроорганизмов не обнаружено. Таким образом, в изучаемой почве удалось выделить только три зоны реакции микробной системы почвы на загрязнение в отличие от четырех зон, характерных для почвы с фоновым содержанием тяжелых металлов. Аналогичные результаты получены и при дополнительном внесении в загрязненный серозем растворимой соли кадмия.
В почве, которая содержит фоновые концентрации тяжелых металлов, большинство микробиологических показателей поддерживается на некотором уровне, в той или иной мере связанном с основными свойствами, определяющими тип почвы. Однако количественные показатели, характеризующие этот уровень, подвержены значительным колебаниям: с другой стороны, именно широкая пространственная и временная вариабельность изменений всех основных микробиологических параметров, обусловленная вполне естественными причинами, является наиболее характерным признаком незагрязненных почв.
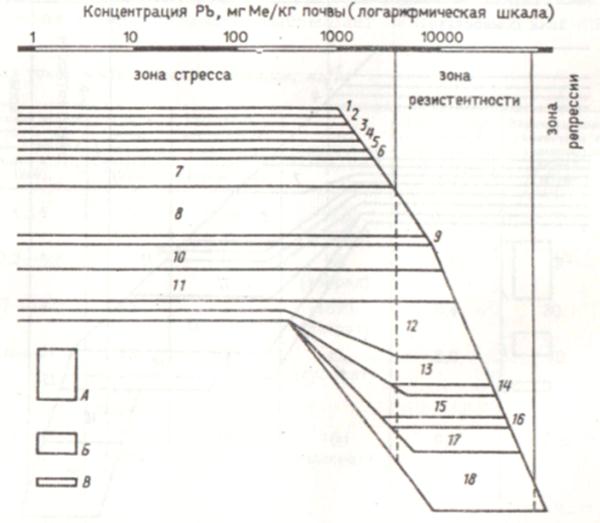
Рис. 21. Влияние свинца (Pb(CH3COO)2) на организацию амилолитического микробного сообщества серозема обыкновенного, загрязненного тяжелыми металлами:
А – доминирующие виды; Б – часто встречающиеся виды; В – редко встречающиеся виды.
1 – Streptomyces sp. cfr. griseus; 2 - Streptomyces sp. cfr. flavus; 3 - Chaetomium anahelicinum; 4 - Monilia geophila; 5 - Aspergillus ustus; 6 - Penecillium funiculosum; 7 - Stysanus stemonites; 8 - Streptomyces sp. cfr. rubro-aurantiacus; 9 - Acremonium charticola; 10 - Monocillium indicum; 11 - Streptomyces sp. cfr. albus; 12 - Stachybotrys alterans; 13 - Coniothyrium macrocladium; 14 – Streptoverticillium sp.; 15 – Penecillium notatum; 16 - Streptomyces sp. cfr. violaceus; 17 - Sepedonium chrysospermum; 18 - Aspergillus flavus
В почвах, где содержание тяжелых металлов превышает фоновое в несколько раз, наиболее заметно изменяются отдельные показатели ферментативной активности. Как правило, несколько возрастает суммарная биомасса амилолитического микробного, сообщества в различных почвах. В той или иной степени изменяются и другие микробиологические показатели. Эти явления свидетельствуют об определенных изменениях в интенсивности микробиологических процессов в почвах. Однако в полевых условиях биохимическая активность почвенных микроорганизмов и большинство других микробиологических характеристик варьируют практически так же сильно, как в незагрязненных почвах.
При дальнейшем увеличении содержания тяжелых металлов в почвах (превышение над фоном до одного порядка) обнаруживается уже достоверное снижение отдельных показателей биохимической активности почвенных микроорганизмов, в том числе в полевых условиях. Характерно, что при указанных концентрациях загрязнителей снижается и вариабельность биохимической активности. Изменения других традиционных показателей микробиологической активности почв, а именно численности различных групп почвенных микроорганизмов и их видового состава, не позволяют обнаружить четкую взаимосвязь с загрязнением почв тяжелыми металлами.
Следует отметить, что при превышении содержания тяжелых металлов в почве на один порядок по сравнению с фоном удается выявить перераспределение степени доминирования членов активно функционирующего в почве амилолитического микробного сообщества.
В почве, содержащей тяжелые металлы в концентрациях, приблизительно на один-два порядка превышающих фоновые, обнаружены достоверные изменения уже целой группы микробиологических показателей. Установлено сокращение богатства видов и видового разнообразия комплекса почвенных микромицетов. Обнаружено абсолютное доминирование по частоте встречаемости ограниченного числа видов или даже одного вида микромицетов. В почве происходит резкое увеличение доли эпифитных дрожжей. Среди различных групп микроорганизмов заметно возрастает количество окрашенных (пигментированных) форм. Это показано и для почвенных микроскопических грибов, и для дрожжей, и для актиномицетов. Увеличение доли пигментированных форм микроорганизмов при загрязнении почв тяжелыми металлами отмечено и для бактерий. В ряде случаев преимущественное развитие в загрязненных почвах получают микроорганизмы· токсинообразователи (Penicillium purpurogenum, Р. funiculosum, Р. janthinellum и некоторые другие).
При превышении содержания тяжелых металлов в загрязненной почве над фоном на три порядка наблюдаются резкие изменения практически всех микробиологических показателей. В целом можно констатировать, что при указанных концентрациях тяжелых металлов в почвах наблюдаются ингибирование и гибель нормальной для незагрязненных почв микробиоты. В то же время обнаружено активное развитие и даже в ряде случаев абсолютное доминирование очень ограниченного числа микроорганизмов, преимущественно микромицетов. По-видимому, эти организмы являются резистентными к тяжелым металлам. В литературе также встречаются сведения о повышенной толерантности некоторых видов почвенных микроорганизмов, в основном микромицетов, к тяжелым металлам. В ряде случаев отмечается, что в биомассе этих микроскопических грибов могут концентрироваться тяжелые металлы.
При концентрациях тяжелых металлов в почвах, превышающих фоновые на четыре и более порядков, обнаруживается катастрофическое снижение микробиологической активности почв, граничащее с полной гибелью микроорганизмов.
Итак, закономерность изменения микробиологических показателей в почве, загрязненной тяжелыми металлами, в градиенте концентраций выглядит следующим образом. При концентрациях этих элементов в почве, равных фоновому содержанию, обнаружено пространственное и временное варьирование всех микробиологических показателей в широком диапазоне, вызванное естественными причинами. При больших концентрациях загрязнителей снижается главным образом активность некоторых почвенных ферментов. Однако вариабельность этого и других показателей, естественная для почв, остается очень значительной. В дальнейшем достоверно снижаются биохимическая активность почв и вариабельность этого показателя, наблюдается перераспределение по степени доминирования активно функционирующих в почве форм микроорганизмов, в то же время многие показатели по-прежнему очень широко варьируют. При дальнейшем увеличении концентраций тяжелых металлов сокращается богатство и разнообразие видов почвенных микромицетов, увеличивается доля пигментированных форм почвенных микромицетов, актиномицетов и дрожжей, эпифитных дрожжей, наблюдается абсолютное доминирование некоторых видов почвенных микроскопических грибов, среди которых могут быть обнаружены токсинообразующие микроорганизм. Затем развиваются преимущественно резистентные к тяжелым металлам формы почвенных микромицетов, а нормальная для незагрязненных почв микробиота ингибируется. При еще больших концентрациях загрязнителей можно зафиксировать практически полную гибель почвенных микроорганизмов.
Поиск по сайту: