АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
Пути радиального транспорта
Корни переносят в надземные органы более 80 % поглощенных ионов. Это перемещение включает радиальный транспорт, загрузку ксилемы и дальний транспорт веществ по сосудам ксилемы. От эпидермиса, где ионы поглощаются из среды, к сосудам ксилемы ионы могут транспортироваться по симпласту и апопласту (рис. 6.19). Соотношение потоков по этим двум компартментам различается для разных ионов и зависит от их концентрации в растворе. При низкой доступности элементов минерального питания, обычной для почвенной среды, большинство ионов, поглощенных в зоне корневых волосков, сразу в ризодерме поступает в симпласт. При высокой концентрации в среде значительная часть ионов (до 50 % от поглощенных) перемещается по апопласту (кальций и бор транслоцируются преимущественно по апопласту). Апопластный радиальный транспорт в корне возможен только до эндодермы, у которой клеточные стенки имеют пояски Каспари (см. подразд. 6.2.1, рис. 6.6, Б и 6.7) и не пропускают воду и растворенные вещества. Поэтому, достигнув эндодермы, все ионы загружаются в симпласт (рис. 6.19).
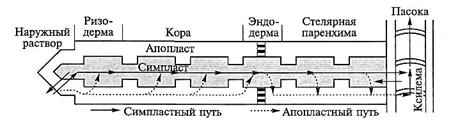
Рис. 6.19. Упрощенная схема путей радиального транспорта ионов по симпласту и апопласту в зоне корневых волосков
Однако непроницаемость клеточных стенок эндодермы не является абсолютной, так как в этой ткани имеются пропускные клетки с тонкими стенками. Развитие боковых корней выше зоны корневых волосков также создает участки, где ионы могут перемещаться из коры в стель по апопласту, поэтому часть ионов и воды может достигать ксилемы, не поступая в симпласт. Симпластная транслокация ионов из клетки в клетку происходит по плазмодесмам, и эффективность перемещения зависит от плотности плазмодесм, пронизывающих клеточную стенку. Число плазмодесм на единице площади поверхности клетки меняется в зависимости от тканевой принадлежности клетки и ориентации стенки (рис. 6.20). В эпидермисе больше всего плазмодесм находится в стенках трихобластов. При этом на тангенциальной стороне плазмодесм больше, чем на радиальной, у трианеи в 3 раза, а у редиса в 1,7 раз (рис. 6.20, А). У безволосковых клеток число плазмодесм в стенках уменьшается по мере удаления от трихобласта.
По направлению от поверхности корня к центру площадь цилиндрической поверхности каждого последующего ряда клеток, а в коре и их число в ряду, уменьшаются (см. рис. 6.7). Для того чтобы скорость симпластного потока не снижалась, число плазмодесм должно возрастать. Действительно, количество и распределение плазмодесм в клеточных стенках меняется таким образом, чтобы обеспечить необходимую скорость радиального перемещения ионов. У корня ячменя плотность плазмодесм на тангенциальном стыке двух клеток коры меньше, чем на стыке кора/эндодерма (рис. 6.20, Б). Но особенно большое число плазмодесм находится на внутренней тангенциальной стороне эндодермы на границе с перициклом. Такое значительное увеличение плотности плазмодесм (двукратное по сравнению с границей кора/эндодерма) необходимо для обеспечения перемещения ионов из двух слившихся потоков — симпластного и апопластного (рис. 6.19). В радиальных стенках перицикла также сосредоточено большое число плазмодесм. Это может быть связано с функцией распределения веществ по периметру стели — ионов, поступающих из коры, и ассимилятов из флоэмы — для более успешного транспорта к местам назначения.
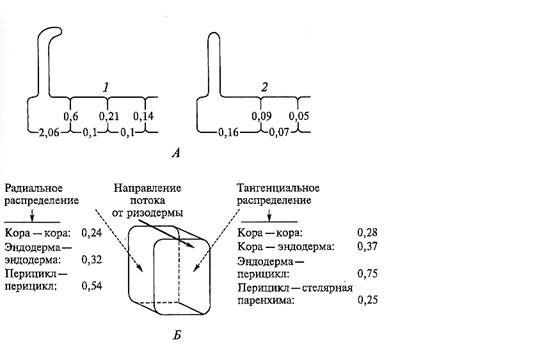
Рис. 6.20. Плотность плазмодесм (число/мкм2) в стенке разных клеток корня:
А — клетки на поперечном срезе эпидермиса: 1 — Trianea bogotensis; 2 — Raphanus sativus (Д. В. Вахмистров и др., 1979; 1981); Б — распределение плазмодесм в радиальных и тангенциальных стенках,на стыках клеток коры, эндодермы и перицикла семинального корня ячменя (Hordeum vulgaris) (по Robards, 1975)
Строение плазмодесмы таково, что она не просто связывает протопласты соседних клеток, но и объединяет через десмотрубочку эндоплазматический ретикулум клеток в единую систему. Наличие подобного отдельного компартмента в симпластном континиуме позволяет объяснить существование потоков веществ, идущих в двух противоположных направлениях: ионов — от поверхности корня к сосудам ксилемы и ассимилятов — из флоэмы в клетки коры и эпидермиса.
Использование того или иного пути для радиального перемещения от поверхности корня до ксилемы зависит от специфики иона, его накопления и распределения в клетке и тканях, а также от его функций. Отдельным компарт-ментом, по которому осуществляется радиальный транспорт, по крайней мере, некоторых ионов, могут быть эндоплазматический ретикулум и десмотрубочки. В качестве теста на радиальное перемещение веществ по эндоплазматическому ретикулуму использовали активность нитратредуктазы (АНР) — цитозольного фермента, который индуцируется собственным субстратом, когда нитрат поступает в клетку через плазмалемму. У растений, выращенных без нитрата, а затем помещенных на среду с низкой концентрацией нитрата, нитратредуктаза индуцируется только в эпидермисе корня, где анион поступает в симпласт (рис. 6.21).
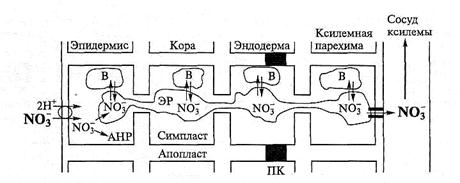
Рис. 6.21. Схема радиального транспорта нитрата в корнях кукурузы при низкой коцентрации NO  (0,2 мМ) в питательном растворе (по Rufty et al., 1986):
(0,2 мМ) в питательном растворе (по Rufty et al., 1986):
ЭР — эндоплазматический ретикулум; В — вакуоль; АНР — активная нитратредуктаза;
ПК — пояски Каспари
Никакой индукции фермента в клетках коры или стели не происходит, хотя значительное количество нитрата обнаруживается в ксилемном экссудате, а, следовательно, нитрат радиально перемещался через корень. Этот транспорт должен идти по симпласту, но очевидно, что потоки нитрата и нитратредуктаза цитозоля каким-то образом изолированы друг от друга. Если корни снабжать нитратом в высокой концентрации (20 мМ), то активная нитратредуктаза обнаруживается в клетках коры и стели, поскольку значительная часть иона движется радиально по апопласту и поступает в клетки всех тканей, индуцируя цитозольную нитратредуктазу (HP). Такие результаты означают, что симпластный транспортный компартмент — это нечто иное, чем вся цитоплазма, а именно эндоплазматический ретикулум. На проростках пшеницы было выяснено, что нитрат, предназначенный для перемещения в побеги, составляет «15 % от общего количества нитрата в корнях. Этот пул, названный «подвижным», представляет собой поток со скоростью обновления «2,5 ч или меньше (см. рис. 6.38).
Возможно, что путь по ЭР используется и для радиального транспорта кальция, концентрация которого в цитоплазме поддерживается чрезвычайно низкой, а в мембране ЭР имеются два типа Са  -АТФаз, с участием которых в люменах ЭР изолируется значительное количество этого иона (см. подразд. 6.3.4.1 и 6.3.4.2). Большее признание получила модель апопластного радиального транспорта кальция по коре и его поступления в симпласт по кальциевому каналу только на уровне клеток эндодермы, откуда он выкачивается в апопласт стели с участием Са -АТФазы плазмалеммы (рис. 6.22).
-АТФаз, с участием которых в люменах ЭР изолируется значительное количество этого иона (см. подразд. 6.3.4.1 и 6.3.4.2). Большее признание получила модель апопластного радиального транспорта кальция по коре и его поступления в симпласт по кальциевому каналу только на уровне клеток эндодермы, откуда он выкачивается в апопласт стели с участием Са -АТФазы плазмалеммы (рис. 6.22).

Рис. 6.22. Транспорт Са2+ из апопласта коры в стель через эндодерму с участием на внешней (коровой) стороне Са2+-каналов и Са2+-АТФаз, выкачивающих катион на внутренней (стелярной) стороне клетки
Клетки эндодермы при этом функционируют аналогично эпителиальным клеткам животных: среда по обе стороны данной ткани различается по ряду параметров (концентрации веществ, метаболической активности и т.д.), а поток Са на входе и выходе обеспечивается разными системами транспорта и регулируется разными механизмами. Известно, что поглощенный анион фосфата в пасоке присутствует также в минеральной форме, перемещаясь исключительно по симпласту. При этом уже в коре фосфор активно включается в состав метаболитов с большими скоростями обновления (см. подразд. 6.3.1.4). Радиальный транспорт фосфора исследовали на растениях кукурузы с использованием методов двойной метки (33Р и 32Р), хроматографии Сахаров, ядерно-магнитного резонанса (ЯМР)31Р, гистохимии фермента глюкозо-6-фосфатазы и с применением ингибиторов. По модели, разработанной на основе результатов, фосфат перемещается через корень в составе глюкозо-6-фосфата (Гл6Ф), а загрузка ксилемы происходит с участием глюкозо-6-фосфатазы, локализованной в плазмалемме клеток ксилемной паренхимы (рис. 6.23).
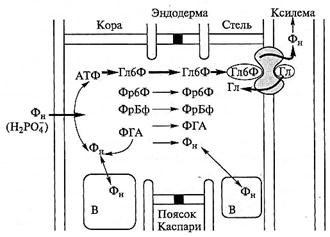
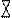 Рис. 6.23. Модель радиального транспорта фосфора и загрузки ксилемы в корнях кукурузы (по Sasaki et al, 1987):
Рис. 6.23. Модель радиального транспорта фосфора и загрузки ксилемы в корнях кукурузы (по Sasaki et al, 1987):
— глюкозо-6-фосфатаза; В — вакуоль
Фермент расщепляет глюкозо-6-фосфат на глюкозу, которая возвращается в клетку, и неорганический фосфат, который переносится в ксилему. Другие фосфорилированные сахара, вероятно, также транспортируются радиально, но их количества незначительны. Быстрое включение в корне поглощенного фосфата в АТФ, а затем в различные органические соединения — факт хорошо известный (см. подразд. 6.3.1.4). Но способ радиального транспорта, специфичный только для фосфата, и особый характер загрузки ксилемы, установленные в результате комплексного исследования на корнях кукурузы, позднее, на других объектах, не были ни подтверждены, ни опровергнуты. В настоящее время идентифицирован транспортер, участвующий в загрузке фосфора в ксилему (см. подразд. 6.3.1.3).
Поиск по сайту: