АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
Індивідуальний розвиток організмів
Розмноження – загальна властивість всіх живих організмів створювати нові покоління особин того ж виду.
Значення розмноження: забезпечує безперервність існування виду, дає можливість збільшити загальну чисельність виду.
Статеве розмноження – спосіб розмноження живих організмів, при якому нові особини утворюються із зиготи, що утворюється в результаті запліднення – злиття статевих клітин – гамет.
Нестатеве розмноження відбувається без участі гамет, без запліднення і в ньому приймає участь один організм.
Партеногенез – розвиток яйцеклітини без запліднення; спостерігається у деяких рослин (кульбаба) та деяких тварин (бджолині трутні, індики).
Поліембріонія:
· у тварин і людини – розвиток з однієї яйцеклітини кількох зародків (однояйцеві близнюки у людини);
· у рослин – утворення кількох зародків в одній насінині.
Партеногенез та поліембріонія з одного боку ведуть до біологічного прогресу, оскільки збільшують чисельність особин виду, а з іншого боку – до біологічного регресу, оскільки при партеногенезі та поліембріонії не відбувається обмін генетичною інформацією і зменшується пристосувальність виду до умов навколишнього середовища
Форми розмноження:
| Форми статевого розмноження | Форми нестатевого розмноження |
| Кон’югація – статевий процес у од-ноклітинних організмів (деяких во-доростей, грибів, більшості інфузорій) при якому від-бувається обмін генетичним матеріалом між двома клітинами. Гермафроди- тизм – здатність одного індивіда ут-ворювати і чоло-вічі і жіночі статеві клітини; характерний для кишковопорожнинних, більшості червів, багатьох молюсків та деяких риб. Статеве розмноження роздільно- статевих видів – в результаті запліднення (зовнішнього чи внутрішнього) та утворення зиготи. | Ділення – утворення двох дочірніх клітин з однієї материнської; клітина може ділитися в будь-якій площині (амеба протей), в вертикальній (евглена зелена) та горизонтальній площині (інфузорія туфелька). Множинний поділ – утворення з однієї материнської клітини великої кількості гаплоїдних клітин (малярійний плазмодій). Спороутворення – процес утворення спор, відбувається у органах нестатевого розмноження спорангіях; характерне для одноклітинних водоростей, вищих спорових рослин, деяких бактерій. Брунькування – поділ материнської клітини при якому утворюються велика і мала дочірні клітини (дріжджі); відбруньковування нових особин, які живуть самостійно (прісноводна гідра), або утворюють колонію (поліпи). Вегетативне розмноження – відбувається з участю відокремлених від материнського організму багатоклітинних частин: вегетативне розмноження рослин – з участю частин вегетативних органів, бульб, цибулин, частин кореневища; фрагментація – розмноження відокремленням певних ділянок тіла (нитчасті зелені водорості, цвілеві гриби, деякі черви) чи спеціальних утворів (лишайники). Клонування – штучний процес, проводиться людиною, полягає в заміні гаплоїдного ядра яйцеклітини на диплоїдне ядро організму який клонують; така яйцеклітина здатна утворювати зародок і плід. |
Статеве розмноження включає в себе процеси:
· гаметогенез – дозрівання і розвиток чоловічих та жіночих статевих клітин (сперматогенез – розвиток чоловічих статевих клітин; оогенез – розвиток жіночих статевих клітин);
· запліднення – процес злиття статевих клітин з утворенням зиготи;
· онтогенез – індивідуальний розвиток організму.
Сперматогенез – процес утворення в чоловічих статевих залозах чоловічих статевих клітин – сперматозоїдів. Сперматогенез проходить в три стадії:
· розмноження в сім’янниках диплоїдних клітин сперматогенної тканини, в результаті якого утворюються сперматоцити, які мають диплоїдний набір хромосом;
· ріст сперматоцитів, який супроводжується синтезом ДНК та добудовуванням другої хроматиди;
· дозрівання сперматоцитів, які діляться мейозом та утворюють гаплоїдні сперматозоїди.
 У сперматозоїдах людини та ссавців хромосомні набори будуть відрізнятися за однією статевою хромосомою: поло-вина сперматозоїдів будуть нести статеву Х-хромосому, інша половина – статеву Y-хромосому.
У сперматозоїдах людини та ссавців хромосомні набори будуть відрізнятися за однією статевою хромосомою: поло-вина сперматозоїдів будуть нести статеву Х-хромосому, інша половина – статеву Y-хромосому.
 Будова сперматозоїда (на прикладі сперматозоїда ссавців). Сперматозоїди дуже малі і рухливі. У ссавців сперматозоїд складається з головки довжиною 5-10 мкм, шийки та хвоста. Загальна довжина сперматозоїда приблизно становить 60 мкм. В головці розміщене ядро з гаплоїдним набором хромосом. Цитоплазми в головці дуже мало. В шийці розміщені мітохондрії, які виробляють енергію для руху сперматозоїда та центріоля, яка забезпечує коливання джгутика, що розміщений вздовж хвоста.
Будова сперматозоїда (на прикладі сперматозоїда ссавців). Сперматозоїди дуже малі і рухливі. У ссавців сперматозоїд складається з головки довжиною 5-10 мкм, шийки та хвоста. Загальна довжина сперматозоїда приблизно становить 60 мкм. В головці розміщене ядро з гаплоїдним набором хромосом. Цитоплазми в головці дуже мало. В шийці розміщені мітохондрії, які виробляють енергію для руху сперматозоїда та центріоля, яка забезпечує коливання джгутика, що розміщений вздовж хвоста.
Оогенез – процес утворення жіночих статевих клітин – яйцеклітин. Оогенез відбувається в жіночих статевих залозах – яєчниках і проходить в три стадії:
· розмноження в яєчниках диплоїдних клітин оогенної тканини, в результаті якого утворюються диплоїдні ооцити;
· ріст ооцитів, який супроводжується синтезом ДНК та добудовуванням другої хроматиди;
· дозрівання ооцитів, які діляться мейозом.
В результаті дозрівання ооцита утворюється одна яйцеклітина і три редукційних (полярних) тільця. Пізніше яйцеклітина бере участь в статевому процесі, а редукційні тільця – відмирають.

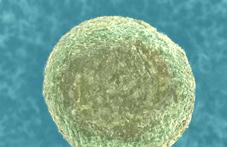 Будова яйцеклітини. Яйцеклітини – малорухливі, переважно великих розмірів, містять запас поживних речовин для розвитку зародка після запліднення. Діаметр яйцеклітини ссавців приблизно 0,1 мм, яйцеклітини риб (ікринки) значно більші, оскільки містять більший запас поживних речовин. Найбільшими є яйцеклітини птахів. Наприклад, яйцеклітина курки разом із запасом поживних речовин (жовтком) має діаметр приблизно 3 см.
Будова яйцеклітини. Яйцеклітини – малорухливі, переважно великих розмірів, містять запас поживних речовин для розвитку зародка після запліднення. Діаметр яйцеклітини ссавців приблизно 0,1 мм, яйцеклітини риб (ікринки) значно більші, оскільки містять більший запас поживних речовин. Найбільшими є яйцеклітини птахів. Наприклад, яйцеклітина курки разом із запасом поживних речовин (жовтком) має діаметр приблизно 3 см.
Форми запліднення:
· у рослин є особливості запліднення: споровим рослинам для запліднення необхідна вода (для руху сперміїв); запліднення квіткових рослин дістало назву подвійного, оскільки один спермій зливається з яйцеклітиною і утворює диплоїдну зиготу, а інший спермій зливається з диплоїдною центральною клітиною і утворює триплоїдну клітину, з якої потім розвивається ендосперм зародка насінини;
· у тварин зовнішнє (відбувається в навколишньому середовищі), внутрішнє (відбувається в організмі самки), штучне (проводить людина); у процесі запліднення відбувається активація амінокислот, проникнення сперматозоїда в яйцеклітину, злиття їхніх ядер.
Онтогенез – індивідуальний розвиток організму, поділяється на дві стадії: ембріональний (зародковий) розвиток та постембріональний (післязародковий розвиток).
Ембріональний розвиток в свою чергу поділяється на стадії:
| Стадії | Особливості |
| Зигота. | Клітина з диплоїдним набором хромосом. |
| Дробіння. | Збільшення кількості клітин в результаті поділу. |
| Морула | Клітини щільно прилягають одна до одної, порожнини немає. |
| Бластула. | Одношаровий зародок. |
| Гаструла. | Двошаровий зародок, що виникає переважно в результаті вгинання (інвагінації) стінки бластули. Зовнішній шар – ектодерма, внутрішній – ентодерма (їх ще називають зародковими листками) |
| Нейрула | Стадія зародкового розвитку хордових тварин і людини, утворення зачатка нервової системи – нервової пластинки і замикання її у нервову трубку. |
| Формування органів та взаємодія частин зародка – органогенез. | Утворюється третій шар клітин – мезодерма, який виростає між екто- та ентодермою. З ектодерми утворюється нервова система, покриви, органи чуття, похідні шкіри, кишечник, зябра (земноводні); з мезодерми – м’язи, скелет, кровоносна, видільна, статева системи, плевра, перикард, сполучнотканинні шари шкіри; з ентодерми – хорда, кишечник, легені, травні залози, плавальний міхур, передня і задня частки гіпофізу, щитовидна залоза і т.д. |
| Вихід зародка із яйцевих оболонок. | Вилуплювання, народження. |
Диференціація клітин – це набуття клітинами здатності до виконання різних функцій, пов’язаних з їх спеціалізацією; процес розвитку клітин у певному напрямку (наприклад, в епітеліальні клітини, нервові клітини та ін.). Диференціація клітин розпочинається у ранньому ембріогенезі і поступово звужує кількість можливих перетворень до однієї або декількох диференційованих форм. Диференційований стан клітин досить стабільний і успадковується їх нащадками. Кінцева диференціація часто супроводжується втратою здатності клітин до розмноження. Наприклад, клітини нервової тканини у ссавців останній раз поділяються в ембріональному та ранньому постембріональному періодах розвитку, а клітини нервових гангліїв дрозофіли – на стадії личинки.
Для пояснення причин диференціації клітин запропоновано дві теорії:
· теорія нерівномірного розподілу спадкового матеріалу серед бластомерів зародка (теорія мозаїчного розвитку);
· теорія однакових генетичних потенцій всіх бластомерів (теорія еквіпотенціальності клітин, або регуляційного типу розвитку).
Прихильником першої теорії був А. Вейсман, який вважав, що мітотичні поділи зиготи є нерівнозначними відносно розподілу генетичного матеріалу. На користь теорії мозаїчного розподілу свідчать наступні факти:
· у кінської аскариди та у деяких інших безхребетних спостерігається втрата хроматину, тобто втрата клітиною частини геному, що не потрібна для даного напрямку диференціації;
· у деяких видів розподіл генетичного матеріалу між статевими та соматичними клітинами здійснюється нерівномірно;
· у багатьох видів (наприклад, у асцидії) спостерігається нерівномірний розподіл ферментів, і-РНК по яйцеклітині, а, отже, по зиготі та бластомерах;
· в процесі розвитку та диференціації клітин можливі незворотні локальні зміни та перебудови генетичного матеріалу (мутації).
На користь теорії регуляційного типу розвитку вказують дані цитогенетики, за якими у переважної більшості організмів каріотипи різних за функцією клітин – однакові (цитофотометричний аналіз вказує на однакову кількість ДНК, а спектральний аналіз показує ідентичність спектрів нуклеотидних послідовностей у різних клітинах). Отже, якщо і є організми, в яких гени і їх продукти нерівномірно розподіляються між клітинами в період дробіння зиготи, то цей феномен не є універсальним. Ембріональні клітини вищих еукаріотів, наприклад, земноводних, людини до стадії восьми бластомерів є генетично ідентичними, тому ембріони цих організмів називають регуляторними. Є і такі організми (наприклад, морський їжак), ембріони яких важко віднести до чисто регуляторних чи мозаїчних.
Відомо, що контроль перших етапів диференціації клітин визначається позаядерними компонентами яйцеклітини, які накопичуються у ній в процесі оогенезу. Деякі з таких компонентів у зародкових клітин дрозофіли ідентифіковані та визначена їх локалізація у яйцеклітині.
Одним із прямих доказів теорії еквіпотенціальності клітин є відомий дослід Дж. Гордона, який трансплантував ядра з диференційованих соматичних клітин (клітин кишкового епітелію пуголовків) в яйцеклітини південноамериканської жаби. Причому, власні ядра незапліднених яйцеклітин вчений руйнував ультрафіолетовим випромінюванням. В окремих випадках із таких штучно сконструйованих яйцеклітин розвивалися дорослі жаби. Отже, можна вважати, що інформація, необхідна для нормального розвитку після багатьох мітотичних поділів не втрачається і може бути використана для повторного процесу розвитку. Такий процес отримав назву клонування, а організми, утворені в такий спосіб називають клонами.
Спроби клонування ссавців довгий час були марними. Однак, у 1997 році група вчених під керівництвом Яна Уілмута у Шотландії шляхом клонування отримали копію вівці за кличкою Доллі.
Подібне клонування сільськогосподарських тварин сьогодні стає буденною справою і має велике практичне значення. Окрім традиційного з використанням незаплідненої яйцеклітини у сільському господарстві здійснюють клонування тварин штучним поділом раннього зародка на частини з наступним вирощуванням з них клонів.
Оскільки ембріональні клітини теплокровних тварин є генетично ідентичними, це можна використати для вирощування тканин та органів необхідних для трансплантації у медицині та ветиренарії. Великого значення набуває технологія вирощування тканин та органів з ізольованих стовбурових клітин, генетично запрограмованих на певний тип диференціювання. Так можна отримати гістосумісний матеріал для трансплантацій без клонування самих організмів.
Клонування людини в технологічному відношенні стало цілком можливим, проте воно стикається з рядом етичних, моральних та психологічних проблем. Гарантувати відтворення особистості клону важко, бо інтелект і все, що з ним пов’язане на 50% залежить від умов життя та виховання.
Постембріональний розвиток: буває прямий (без метаморфозу) та непрямий (з метаморфозом).
За прямого розвитку щойно народжена тварина загалом нагадує дорослу особину. Прямий розвиток відбувається внаслідок ембріонізації – періоду, коли зародок живиться за рахунок материнського організму (плацентарні ссавці), або запасними поживними речовинами яйця (плазуни, птахи).
За непрямого розвитку з яйця виходить личинка, що зовсім не схожа на дорослих особин ні будовою ні способом живлення, а часом для неї характерне ще й інше середовище життя. Личинка живиться, росте та розвивається (деякі види можуть на личинковій стадії розмножуватись), а пізніше у її організмі відбуваються глибокі структурні зміни, завдяки яким личинка перетворюється у дорослу комаху. Непрямий розвиток може відбуватися з повним та неповним перетворенням:
Неповне перетворення: яйце → личинка, схожа на дорослу особину → доросла особина.
Повне перетворення: я йце → личинка, що несхожа на дорослу особину → лялечка → доросла особина.
Ріст – кількісні зміни в організмі, один з проявів індивідуального розвитку організму, одна з ознак живого, пов’язаний з збільшенням маси, лінійних розмірів та об’єму організму, супроводжується диференціацією клітин і таким чином пов’язаний з розвитком. Ріст буває необмежений – відбувається протягом всього життя організму (рослини) та обмежений – відбувається до певного періоду індивідуального розвитку організму (людина).
Регенерація – здатність організмів відновлювати втрачені чи пошкоджені органи. Процеси регенерації у тварин регулюються: спадково, гормонами, нейрогормонами, імунною системою, а також міжклітинними та між тканинними взаємодіями.
Регенерація буває:
· репаративна – відновлення втрачених структур (загоєння ран, відновлення ящіркою втраченого хвоста);
· фізіологічна – відновлення зношених структур (клітин крові, клітин епідермісу шкіри та ін.)
Життєвий цикл (цикл розвитку) – сукупність фаз розвитку, при завершенні яких організм досягає статевої зрілості та досягає здатності до розмноження.
Життєвий цикл організмівбуває простим або складним:
· за простого життєвого циклу всі послідовні покоління не відрізняються одне від одного;
· за складного життєвого циклу відбувається чергування поколінь: статевого, переважно гаплоїдного покоління (гаметофіту) та нестатевого диплоїдного покоління (спорофіту).
Біологічне значення чергування поколінь у деяких організмів полягає у кращому пристосуванні до умов середовища, що змінюється: статеве розмноження забезпечує існування виду в нестабільних умовах середовища, а нестатеве – дозволяє швидко збільшити чисельність виду в стабільних, сприятливих умовах середовища.
Поиск по сайту: