АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
Антропогенных воздействий
Пусть x и x*- величина биоиндикационного признака и его нормальное значение соответственно; z = (x - x*) - уклонение значения признака от нормы; n(z) - число особей в популяции, имеющих уклонение индикационного признака от нормы, равное z; A(z) - детерминированная внешним воздействием скорость уклонения признака от нормы; B - интенсивность случайных уклонений признака от нормы. Тогда величина потока особей популяции биоиндикатора, перемещающихся по шкале величин уклонений индикационного признака от нормы, равна (Алексеев,1997)
S = A(z)n(z) - B(dn/dz).
Изменение во времени распределения особей биоиндикаторов по величине уклонения индикационного признака от нормы должно подчиняться уравнению Фоккера - Планка (Хакен,1980):
(dn/dt) + dS/dz = 0,
откуда для стационарного случая (dn/dt = 0) при дополнительном реалистичном условии n Þ 0 при z Þ àà получаем, что S=0 или
dn/dz = [A(z)/B] n(z).
Из последнего выражения следует, что стационарное распределение особей индикаторов в популяции по величине уклонения биоиндикационного признака от нормы определяется следующим выражением:
n(z) = n* exp[-V(z)/B], (55)
где n* - постоянная, равная числу (доле) особей в популяции, имеющих значение индикационного признака, равное норме, z = 0; V(z) = -òA(z)dz, так, что A(z) = - dV(z)/dz.
Функция V(z) представляет собой потенциал устойчивости популяции биоиндикаторов к внешнему антропогенному воздействию, так как ее убыль равна величине скорости уклонения индикационного признака особей от нормы.
Введем системно-популяционные характеристики. Определим значение константы n* из условия нормировки:
àà
ò n(z)dz = 1,
-àà
откуда имеем
àà àà
ò n(z)dz = n* ò exp[-V(z)/B]dz = 1
-àà -àà
и àà
n* = 1/Z, Z = òexp[-V(z)/B]dz,
-àà
где Z - статистическая сумма распределения. Определим характеристическую функцию популяции биоиндикаторов F следующим образом:
F = -Bln(Z) = Bln(n*).
С ее помощью распределение особей биоиндикаторов по величине уклонения индикационного признака от нормы (55) можно представить в следующем каноническом виде:
n(z) = exp[(F - V(z))/B]. (56)
Функцию F можно интерпретировать как нормальный потенциал популяции биоиндикаторов, так как она определяется численностью особей, для которых значение индикационного признака равно норме (z = 0). Рассмотрим энтропию распределения особей в популяции биоиндикаторов по величине уклонения индикационного признака от нормы:
àà àà
H = - ò n(z)ln(n(z))dz = -ò exp[(F - V(z))/B][(F - V(z))/B]dz.
-àà -àà
Нетрудно показать, что
H = - (F - R)/B,
àà
где R = ò V(z)n(z)dz - средний потенциал устойчивости популяции
-àà
биоиндикаторов к внешнему воздействию. Средний потенциал устойчивости популяции связан с нормальным потенциалом следующей зависимостью:
R = F + BH.
Произведение ВН можно интерпретировать как внутренний потенциал устойчивости популяции биоиндикаторов, определяемый ее внутренней структурой.
Для применения на практике развиваемой теории популяционной биоиндикации необходимо определить вид функции A(z) - детерминированной внешним воздействием скорости уклонения индикационного признака особей от нормы. Исходя из общебиологических соображений разумно предположить, что вид функции будет типа кривой с насыщением, т.е. величина скорости уклонения индикационного признака от нормы будет убывать со временем пропорционально величине такого уклонения:
A(z) = dz/dt = E - z.
Выбор такого вида функции предполагает наличие в популяции механизмов самоорганизации и саморегуляции, обеспечивающих компенсацию уклонений индикационного признака особей от нормы. Величина E характеризует интенсивность детерминированного внешнего воздействия, вызывающего уклонение признака от нормы. Если E < 0, то внешнее воздействие снижает значение признака ниже нормы, т.е. имеет травмирующий характер, если E > 0, то воздействие повышает значение признака выше нормы, т.е. имеет стимулирующий характер.
Принимая во внимание предлагаемый вид функции A(z), получим следующее выражение для распределения особей в популяции по величине уклонения индикационного признака от нормы:
n(z) = n*exp[-(1/B)(z2/2 +Ez)]. (57)
Это распределение нормального (Гауссова) типа. В нормированном виде его можно записать следующим образом:
n(z) = (1/2pB)-1/2 exp[-(1/2B)(z + E)2]. (58)
Сравнение с нормальным распределением в обычной записи позволяет установить следующие зависимости между статистическими и синергетическими характеристиками популяции биоиндикатора:
z* = -E, s2 = B, (59)
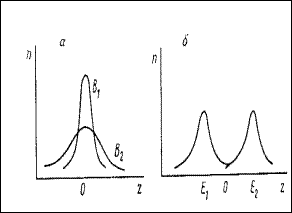
где z* и s2 - среднее значение и дисперсия нормального распределения. Таким образом, характеристика детерминированной скорости уклонения индикационного признака в популяции от нормы определяет среднее значение такого уклонения, а интенсивность случайного отклонения от нормы - дисперсию уклонений вокруг среднего. Вид распределения (57) при различных сочетаниях его параметров представлен на рис.21.
Выразим через параметры распределения (57) системные характеристики популяции биоиндикаторов: нормальный потенциал (F), средний потенциал устойчивости (R) и энтропию (H):
нормальный потенциал F = - (E2/2) - Bln((2pB)-1/2),
средний потенциал, R = (B/2) - (E2/2),
энтропия распределения, H = ln((2pB)-1/2) + 1/2.
|

|
Рис. 21. Формы распределения деревьев по величине уклонения индикационного признака от нормы при разном соотношении параметров В и Е: а-В1<В2; б-Е1<0, Е2>0, В-const. По оси абцисс - величина отклонения индикационного признака от нормы (z), по оси ординат - число деревьев (n) (по Алексеев, 1997)
Рис. 22. Распределение деревьев по величине отклонения индикационного признака от нормы на пробной площади А в 1976 и 1986 гг. По осям то же, что на рис.21
Оценку изменений в состоянии популяции организмов - индикаторов, находящихся под внешним антропогенным воздействием, можно осуществить по изменениям ее системных характеристик и их баланса:
DR = DF + D(BH). (60)
Отметим, что такие системные характеристики популяции биоиндикаторов как средний потенциал устойчивости R и нормальный потенциал F будут одинаково изменяться как в случае травмируюшего, так и стимулируюшего внешнего воздействия. Принципиальным здесь является уклонение у большого числа особей в популяции индикационного признака от нормы, причем неважно, в какую сторону. Однако с экологической точки зрения различие между положительным и отрицательным внешним воздействием имеет важное значение. Важно также достоверно выделить на фоне случайных изменений индикационного признака в популяции детерминированную внешним воздействием тенденцию. Для этого будем использовать показатель E/(B1/2) (McArthur,1965,1966; Theoretical Ecology...,1976; Пианка,1981; Джиллер,1988). Величина E характеризует детерминированную внешним воздействием скорость уклонения индикационного признака у особей от нормы, ее знак показывает положительный или отрицательный характер этого воздействия. Величина B характеризует интенсивность случайных уклонений индикационного признака от нормы в данной популяции в данное время. Корень из этой величины есть стандартное уклонение признака от нормы. Исходя из вышеизложенного при популяционной биоиндикации внешних антропогенных воздействий возможны следующие четыре варианта:
1. Если E/(B1/2) > 1, то имеет место достоверное положительное воздействие, при котором детерминированное уклонение выходит за рамки стандартного случайного.
2. Если 0 < E/(B1/2) < 1, то имеет место недостоверное положительное внешнее воздействие, детерминированное уклонение от нормы находится в пределах стандартного для данной популяции случайного отклонения.
3. Если -1 < E/(B1/2) < 0, то имеет место недостоверное отрицательное внешнее воздействие на популяцию биоиндикаторов, его детерминированная составляющая в пределах стандартного отклонения.
4. Если E/(B1/2) < -1, то имеет место достоверное отрицательное воздействие на популяцию биоиндикаторов. Чем больше по модулю величина E/(B1/2) превосходит единицу, тем более достоверно наличие внешнего антропогенного воздействия.
Разработанная теория популяционной биоиндикации была применена для анализа влияния загрязнения атмосферы на рост насаждений ели европейской и сосны обыкновенной, в качестве биоиндикационного признака использовались индексы радиального прироста деревьев, сгруппированные по величине в 11 классов (Алексеев, 1997):
Поиск по сайту: