АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
Сопряжение обмена липидов и аминокислот. Сопряжение обмена углеводов, липидов и аминокислот на уровне образования ацетил-КоА, и на уровне цикла Кребса
Гормоны гипоталамуса и нейрогипофиза, их химическая природа и влияние на обмен веществ.
Оценить состояние больного по следующим данным анализа его крови: общая активность ЛДГ повышена, коэффициент АСТ/АЛТ – 0,8, общий билирубин – 55 мкмоль/л, непрямой билирубин – 30мкмоль/л, прямой билирубин – 25 мкмоль/л, количество альбуминов снижено, концентрация альфа-2- и бета-глобулинов повышена.
1. Взаимосвязь обменов осуществляется благодаря интегрирующим системам: нервной, эндокринной, сосудистой.
Взаимосвязь обеспечивается различными уровнями: информационный уровень, структурный уровень, общее энергетическое обеспечение, на уровне общих метаболитов, на уровне Ц.Т.К.
Информационный уровень взаимосвязи. В геноме клеток заложена информация о структуре и функциональной активности различных белков, принимающих участие в структурной и динамической организации живых систем.
Структурный уровень взаимосвязи. Мембранный аппарат клеток, рибосомы.
Общее энергетическое обеспечение. АТФ – универсальная энергетическая валюта, образующаяся при окислении углеводов, жиров, аминокислот. НАДФН2 – основной донор электронов в восстановительных реакциях биосинтеза. Восстановительные эквиваленты, накапливаемые в ходе катаболизма в клетке в виде восстановительных форм НАДФН2 используются в восстановительных реакциях клеточного анаболизма, связывая таким образом, катаболические и анаболические процессы в единую систему. Синтез одного соединения (жира) происходит за счёт катаболизма другого (глюкозы).
Взаимосвязь на уровне общих метаболитов. Центральные метаболиты: ацетил-КоА, ПВК, ЩУК, ФГА.
Ацетил-КоА образуется при окислительном декарбоксилировании ПВК, при β-окислении жирных кислот, из аминокислот. 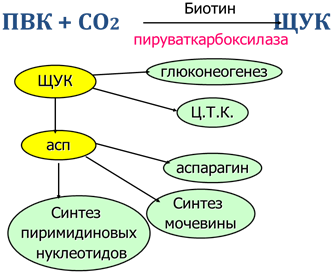
Взаимосвязь углеводного и липидного обменов осуществляется через - ацетил-КоА, ФГА, НАДФН2 из пентозного цикла идёт на синтез жирных кислот, ЩУК нужен для работы Ц.Т.К.
Жиры сгорают в пламени углеводов. При избытке в пище углеводов возникает ожирение. ФГА и ацетил-КоА– источники глицерина и жирных кислот. При спячке у животных происходит образование углеводов из жиров. Взаимосвязь белкового и жирового обменов на уровне - ПВК, ацетил-КоА, кетоновых тел. Из белков осуществляется синтез жира.
Взаимосвязь углеводного и белкового обменов на уровне - ПВК, ЩУК, ацетил-КоА, пентоз, глюкопластичных и кетопластичных аминокислот. Углеродные скелеты АМК вступают в Ц.Т.К. Возможно образование углеводов из белков (ГНГ) и белков из углеводов (из ПВК образуется аланин).
Сопряжение на уровне Ц.Т.К. - углеродные скелеты АМК включаются в Ц.Т.К, все кислоты Ц.Т.К. превращаются в ЩУК (ГНГ), взаимосвязь Ц.Т.К. с синтезом мочевины через фумарат, асп, СО2.
Ц.Т.К. обеспечивает энергией все обмены. Взаимосвязь обмена веществ на уровне цикла Кребса. Связь липидного обмена с Ц.Т.К. осуществляется через: цитрат, - активатор ацетил-КоА-карбоксилазы, - перенос в цитоплазму ацетил-КоА, Сукцинил-КоА.
Жирные кислоты с нечётным числом углеродных атомов через пропионил-КоА превращаются в сукцинил-КоА, СО2, малат. Малик-реакция – источник образования НАДФН2.
2. Гормоны гипоталамуса (релизинг-факторы) - соматолиберин, пролактолиберин, тиролиберин, кортиколиберин, люлиберин, меланолиберин, фоллилиберин, соматостатин, меланостатин, пролактостатин.
Соматостатин. Он оказывает широкий спектр биологического действия; в частности, показано ингибирующее действие на синтез гормона роста в аденогипофизе, а также прямое тормозящее действие его на биосинтез инсулина и глюкагона в β- и α-клетках островков Лангерганса.
Соматолиберин стимулирует синтез и секрецию гормона роста гипофиза соматотропина.
Меланолиберин оказывает стимулирующее действие, а меланостатин, напротив, ингибирующее действие на синтез и секрецию меланотропина в передней доле гипофиза.
Местом синтеза гипоталамических гормонов, вероятнее всего, являются нервные окончания – синаптосомы гипоталамуса, поскольку именно там отмечена наибольшая концентрация гормонов и биогенных аминов. Последние рассматриваются наряду с гормонами периферических желез внутренней секреции, действующих по принципу обратной связи, в качестве основных регуляторов секреции и синтеза гормонов гипоталамуса.
Механизм биосинтеза тиролиберина, осуществляющегося, скорее всего, нерибосомальным путем, включает участие SH-содержащей синтетазы или комплекса ферментов, катализирующих циклизацию глутаминовой кислоты в пироглутаминовую, образование пептидной связи и амидирование проли-на в присутствии глутамина. Существование подобного механизма биосинтеза с участием соответствующих синтетаз допускается также в отношении гонадолиберина и соматолиберина.
Гипоталамические гормоны непосредственно влияют на секрецию (точнее, освобождение) «готовых» гормонов и биосинтез этих гормонов de novo. Доказано, что цАМФ участвует в передаче гормонального сигнала. Показано существование в плазматических мембранах клеток гипофиза специфических аденогипофизарных рецепторов, с которыми связываются гормоны гипоталамуса, после чего через систему аденилатциклазы и мембранных комплексов Са2+–АТФ и Mg2+–АТФ освобождаются ионы Са2+ и цАМФ; последний действует как на освобождение, так и на синтез соответствующего гормона гипофиза путем активирования протеинкиназы.
Гормоны задней доли гипофиза. Вазопрессин и окситоцин синтезируются внейронах гипоталамуса, связываются с белками нейрофизинами и транспортируются в нейросекреторные гранулы гипоталамуса, затем вдоль аксона в заднюю долю гипофиза, где происходит пострибосомальная достройка.
Вазопрессин - стимулятор аденилатциклазы: цАМФ образуется в мембране эпителия почечных канальцев, в результате повышается проницаемость для воды, повышает артериальное давление из-за стимуляции сокращения гладкой мускулатуры сосудов, способствует уменьшению диуреза из-за воздействия на канальцевый аппарат нефрона, повышения реабсорбции воды.
Несахарный диабет возникает из-за нарушения: синтеза, транспорта, секреции вазопрессина. При заболевании с мочой теряется до 40 л воды в сутки, возникает жажда. Несахарный диабет бывает при атрофии задней доли гипофиза.
Синдром Пархана возникает из-за повышенной секреции вазопрессина, усиливается реабсорбция воды в почках, появляются отёки.
Окситоцин - стимулирует сокращения гладкой мускулатуры матки, гладких мышц кишечника, уретры, стимулирует сокращение мышц вокруг альвеол молочных желёз, способствуя молокоотдаче. Окситоциназа разрушает гормон. При родах её активность падает в 100 раз.
3. Ответ. ↑ ЛДГ – болезнь печени, сердца, легких.
Коэффициент Де Ветиса < 1 – забол. печени
↑ непр. и прям. Билирубина – паренхиматозная желтуха.
↑ концентрации белков острой фазы – острое воспаление.
Билет 40.
Поиск по сайту: