АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
Класифікація типів зародкового міхура (зм)
Відомо декілька класифікацій типів розвитку зародкових міхурів. В основу сучасних класифікацій прийнято:
1) число мегаспор, що беруть участь в утворенні зародкового міхура (моно-,
бі-, тетраспоричні зародкові міхури);
2) кількість ділень ядра, починаючи від мегаспори і завершуючи зрілим зародковим міхуром;
3) поляризацію, тобто остаточний розподіл ядер у зрілому зародковому міхурі.
Найбільш детальна (16 типів зародкових міхурів) класифікація, яка відображує філогенію зародковог міхура у покритонасінних запропонована І.Д. Романовим (1971). Рис.
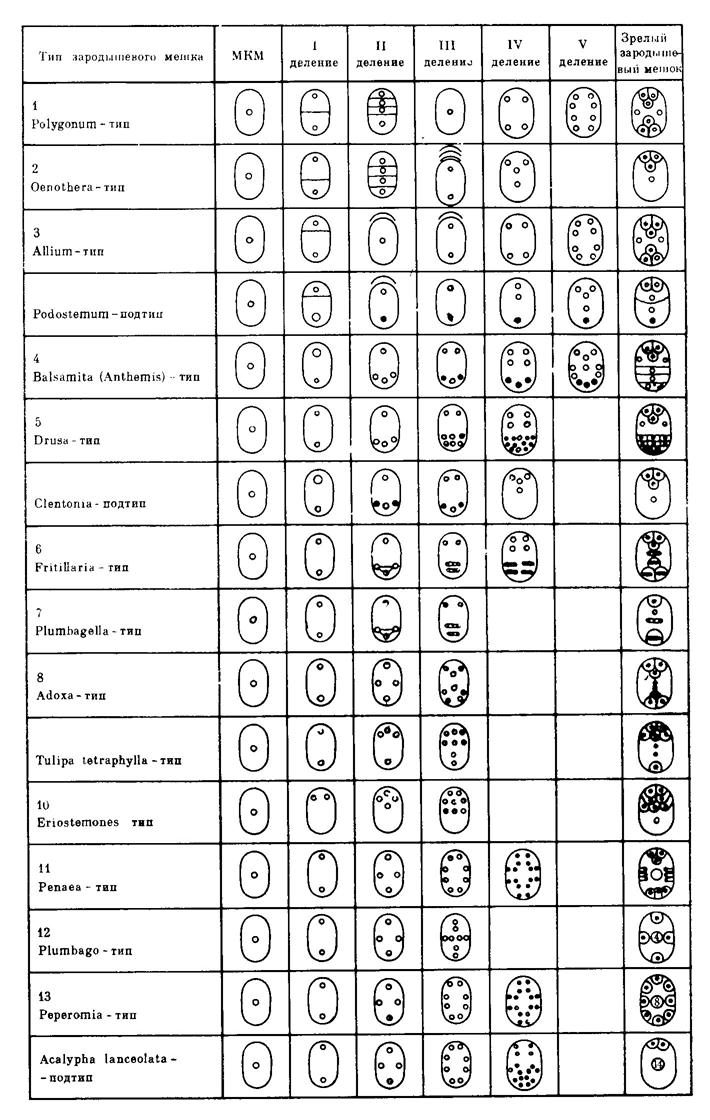
Рис. Класифіуація типів зародкового міхура (Романов,1971).
Залежно від числа мегаспор, що беруть участь в утворенні, зародкові міхури ділять на три типи:
1) односпорові - моноспоричні;
2) двоспорові – біспоричні.
3) чотириспорові – тетраспоричні.
Моноспоріальний тип описав ще Страсбургер у Polygonum divaricatum L. (1879). (Звідси й назва - Polygonum- тип). За цього типу в зародковий мішок розвивається лише одна халазальна макроспора (=мегаспора), три інші відмирають.
При розвитку восьмиядерного зародкового мішка із халазальної макроспори (Polygonum-тип) відбувається п’ять поділів: перше і друге ділення мейозом і три мітозом. Після другого ділення мейозу нижня (халазальна) мегаспора сильно розростається, що супроводжується вакуолізацією і поляризацією обміну речовин: синтез білків і нуклеїнових кислот у верхній частині клітини відбувається більш інтенсивно, ніж у нижній.
Моноспоричні зародкові мішки формуються і за Oenothera–типу розвитку, за якого зародковий мішок містить лише чотири ядра і також утворюється з мікропілярної макроспори. Проте весь цикл розвитку складається з чотирьох поділів, у результаті яких утворюється яйцевий апарат із трьох клітин і верхнє полярне ядро. Антиподи відсутні.
За біспоріального Allium- типу формування зародкового мішка відбувається з нижньої - халазальної - двоядерної мегаспори (гаплоїдної), верхня швидко дегенерує.
При Allium – типі розвитку зародкового міхура відбувається чотири поділи (замість
п’яти як це у Polygonum-типу), відраховуючи від першого ділення материнської клітини спор до зрілого зародкового міхура. Скорочення на одне ділення обумовлено тим, що перше ділення при розвитку зародкового міхура із двох мегаспор є одночасно другим діленням мейозу.
Морфологія зрілих зародкових міхурів Allium- і Polygonum-типу дуже подібна, і в тому і другому випадках утворюється двополюсний восьмиядерний зародковий міхур.
Отже, онтогенез біспоріального зародкового міхура проходить швидше, ніж моноспоріального, і зародковий міхур утворюється двома мегаспорами. Біспоріальний тип зародкового міхура у проліски, цибулі, сусака.
Чотириспорові (тетраспорові) зародкові міхури вирізняються широкою різноманітністю будови. До цієї групи належить десять типів розвитку зародкових мішків. При їх утворенні ні в першому, ні в другому діленнімейозу цитокінез не відбуається. Загальною ознакою цієї групи є участь усіх ядер мегаспор у розвитку і формуванні зародковогоміхура.
Під час мейозу ні при першому, ні при другому діленні цитокінез не відбувається. Зародкові міхури цієї групи суттєво відрізняються як за числом ядер і клітин, так і за їхнім розміщенням у зародковому міхурі.
В літературі є чимало повідомлень про різні аномалії зародкового міхура, яйцевого апарату, антипод. викликаними різкою зміною зовнішніх умов, віддаленою гібридизацією, радіацією тощо.
Аналіз перебігу розвитку зародкового міхура рослин показує, що чим менше часу затрачається на мегаспорогенез і мегагаметогенез завдяки скороченню кількості поділів клітин, тим прогресивніша ця форма. У голонасінних жіночий гаметофіт формується майже рік, при цьому клітини багаторазово діляться. Покритонасінні на цей процес витрачають кілька днів і навіть годин з 5-6 поділами клітин. Тобто. спостегігається значний прогрес, бо енергії на той самий процес витрачається набагато менше.
Зародковий мішок у покритонасінних має свій незалежний шлях розвитку, і є якісно новим явищем, що притаманне лише даній групі рослин. Серд інших груп вищих рослин (архегоніальні, голонасінні) він не зустрічається. А тому усі намагання встановити взаємозв’язок між будовою жіночого гаметофіту голонасінних і зародковим мішком покритонасінних поки що не стали успішними.
СР:
Будова гінецею;
Розвиток нуцелуса, археспорію, мегаспорогенез та мегагаметогенез;
Формування зародкового міхура;
Відхилення від Polygonum-типу зародкового міхура;
Варіювання будови яйцевого, aнтиподного апаратів і вторинного ядра;
Класифікація типів жіночого гаметофіту.
Порушення нормального розвитку і будови жіночого гаметофіту.
Методи вивчення структури, розвитку зародковогоміхура.
Л- ура: Стеблянко М.І., Гончаров К.Д., Закорко Н.Г. Ботаніка К: Вища шк. 1995. с.275-308.
Поддубная – Арнольди В.А.Цитоэмбриолог. покрытосем - х раст. 1976. с.143-210
Устинова Е И.Эмбриолог. покрытосем - х раст. с основами цитолог. М 1965. с.90-110
Розділ 5
БІОЛОГІЯ ЗАПИЛЕННЯ
Анатомічні і морфологічні, біохімічні та фізіологічні особливості будови пилкових зерен і приймочки маточки. Екологічні аспекти запилення. Адаптації рослин до способу запилення. Причини порушення нормального перебігу процесів запилення та запліднення.
і закрите запилення, гейтоногамія. Поняття сумісності, несумісності, самонесумісності. Структурні і генетичні механізми несумісності, її біологічна значимість та врахування практичній діяльності людини.
Способи подолання несумісності та стерильності.
Перенесення пилку на приймочки.
Дихогамія, протандрія, протогінія і гетеростилія.
Взаємодія пилкове зерно - приймочка маточки, пилкова трубка – стовпчик маточки.
Проростання пилкового зерна і швидкість росту пилкової трубки за різних способів і умов
Поиск по сайту: