АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
Древние, старые и межуточные корковые формации
В отличие от новой коры (пеосоrtех), составляющей большую часть мозгового плаща у человека, древние (paleocortex) и старые (archicortex) компоненты ее представлены только в отдельных участках медиальной и базальной поверхностей полушарий. Эти отделы появляются ранее в филогенезе позвоночных и более примитивны по своей внутренней структуре. Архи- и палеокортекс, вместе с прилегающими к ним переходными образованиями межуточной коры (mesocortex), не имеют шестислойного строения, которое обязательно проявляется на определенной стадии онтогенеза в неокортексе. Это позволяет классифицировать неокортекс как гомогенетическую кору, а все остальные ее отделы – как гетерогенетические.
Древняя кора слабо обособлена от подлежащих подкорковых образований. К ней относятся латеральная обонятельная извилина, называемая также препериформной областью, передняя часть парагиппокампальной извилины, которая вместе с крючком и подлежащим миндалевидным комплексом рассматривается как периформная область, и, наконец, медиальная обонятельная область, в которую включаются обонятельный бугорок, диагональная извилина, подмозолистая извилина, парольфакторная зона, а также прозрачная перегородка (рис. 7.12 и 7.13).
Структура палеокортекса представляет собой скопление многочисленных, не имеющих отчетливо выраженного разделения на отдельные слои, клеточных островков. Основные афференты подходят к ним через наружный зональный слой.
Окружающие древнюю кору в местах перехода к новой коре участки (peripaleocortex) включают 3 слоя. Наружный слой образован островковыми клетками, а два других включают полиморфные нейроны, разделенные пучками волокон.
Основными структурами старой коры являются гиппокамп и зубчатая извилина. Гиппокамп (морской конек) представляет собой ввернутый внутрь полости нижнего рога бокового желудочка участок в стенке и дне глубокой гиппокампальной борозды. Зубчатая извилина является самой крайней структурой мозгового плаща, которая сращена с гиппокампом и заворачивается над ним (рис. 7.13 и 7.15).

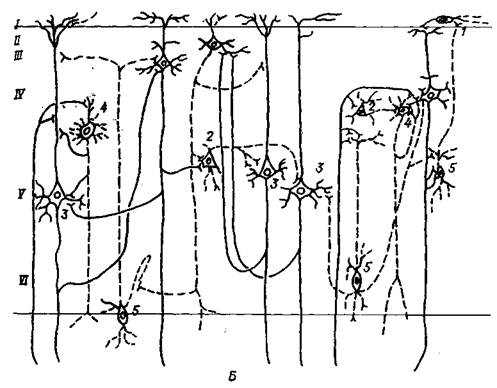
Рис. 7.15. Схемы нейронной организации гиппокампа (А) и участка новой коры (Б):
А: 1 – аммонов рог; 2 – бахрома; 3 – альвеус; 4 – слой пирамидных клеток; 5 – радиальный слой; 6 – молекулярный слой; 7 – слой полиморфных клеток; 8 – субикулум; 9 – энторинальная кора; 10 – молекулярный, гранулярный и полиморфный слои зубчатой извилины; 11 – волокна свода; 12 – аф-ференты от субикулума; 13 – афференты от прозрачной перегородки; 14 – корковые энторинальные афференты; Б: 1 – горизонтальная клетка; 2 – гранулярная клетка; 3 – пирамидная клетка; 4 – звездчатая клетка; 5 – веретеновидная клетка; I – VI – слои коры
В гиппокампе и зубчатой извилине четко различаются два клеточных слоя. Наружный слой гиппокампа (stratum pyramidum) образован довольно крупными пирамидными нейронами. Ему гомологичен гранулярный слой (stratum granulosum) зубчатой извилины. Внутренний клеточный слой аммонова рога образован полуморфными нейронами (stratum polimorpha) (рис. 17.15, А).
Снаружи гиппокамп покрыт белым веществом(alveus), сильно вдающимся в полость нижнего рога бокового желудочка, и включающем аксоны пирамидных клеток гиппокампа. Проходя затем на медиальную сторону зубчатой фасции, эти миелинизированные волокна образуют бахромку (fimbria). Далее они переходят в свод, направляясь под мозолистым телом частью к ядрам прозрачной перегородки, а основной массой к мамиллярному телу и частично в другие отделы гипоталамуса с той же стороны. Некоторые волокна заканчиваются в противоположном гиппокампе, проходя через комиссуру свода.
Апикальные дендриты клеток гиппокампа тянутся без разветвления кнаружи, образуя особый радиальный cлой (stratum radiatum). Далее они интенсивно ветвятся с образованием молекулярного cлоя (stratum moleculare), который срастается с зональным молекулярным слоем (stratum zonale) зубчатой извилины. Таким образом, в зубчатой извилине можно насчитать всего 3, а в гиппокампе – до 5 слоев.
Основная часть афферентных окончаний в гиппокампе располагается на тонких ветвлениях дендритов, густо покрытых шипиками. Лишь очень незначительное число синапсов контактирует с телами пирамидных клеток. Базальные дендриты пирамид гиппокампа ориентированы в направлении, противоположном апикальным дендритам. Тела этих нейронов вначале располагаются в один ряд, но постепенно, по мере перехода к мезокортексу, их слой становится шире. В соответствии с цитоархитектоническими различиями в витке, который делает гиппокамп, различают пять полей (Н5 – Н1). Последнее поле (H1) граничит с периархикортексом – промежуточной областью между старой и новой корой. Здесь происходит перекрытие элементов, свойственных неокортексу и гиппокампу с постепенным нарастанием наружной зоны неокортикальных структур по мере удаления от гиппокампа. В соответствии с цитоархитектоническими особенностями периархикортикальная область делится на ряд полей. Ближайший к гиппокампу участок переходной коры – субикулум (subiculum) расположен в гиппокампальной извилине. Он характеризуется широким зональным слоем, нечетко отделенным от диффузно расположенных клеток средней величины, образующих широкую корковую пластинку (рис. 7.15, А).
В поле Н1 и особенно Н2 гиппокампа корковая пластинка значительно уже и имеет более плотное расположение клеток, чем в субикулуме. В поле Н3 – Н5 клеточная пластинка становится все более разреженной.
Корковые структуры, характерные для гиппокампа и зубчатой фасции, по ходу ножки свода вверх постепенно редуцируются и в таком виде присутствуют в сером покрове мозолистого тела. Эта промежуточная структура между старой и древней корой включает слой небольших пирамидных клеток, образующих в латеральных участках серого покрова от 5–6 рядов в задней части до 1–2 рядов в передней и средней частях.
Зубчатая фасция примыкает к зональному слою субикулума и полям аммонова рога. Она составлена компактной мелкоклеточной корковой пластинкой. В зубчатой извилине основной клеточный слой (stratum granulare) состоит из плотно расположенных овальных и мультиполярных мелких зернистых нейронов. В расположенном ниже внутреннем слое концентрация полиморфных клеток резко снижается и они приобретают диффузное распределение.
Дендриты гранулярных и полиморфных клеток направляются к гиппокампальным пирамидам, а их аксоны подходят в виде мшистых волокон к дендритам клеток пирамидного слоя гиппокампа. Старая кора отделяется от неокортекса переходными зонами, включающими пресубикулярную и энторинальную области. Корковая пластинка этих областей делится на наружный, средний и внутренний слои, причем в двух первых отчетливо выявляются светлые прослойки, а последний переходит в корковую пластинку архикортекса. Особенно сложное расслоение наблюдается в энторинальной области, которая снаружи ограничивается задней ринальной бороздой, а изнутри немного не доходит до гиппокампальной борозды.
Пресубикулярная область занимает верхнюю часть наружной стенки гиппокамповой борозды, перешеек, верхнюю стенку борозды мозолистого тела и парольфакторное поле.
Препериформная область образована примитивно построенной корковой пластинкой, характеризующейся широким зональным слоем. Часть периформной области, выходящая на свободную поверхность, представлена более широкой корковой пластинкой, состоящей из средне-, крупно- и мелкоклеточных зон. В обонятельном бугорке зональный слой выражен слабо и его корковая пластинка отличается густым распределением мелких клеток.
Прозрачная перегородка в верхней части содержит лишь зональный слой и практически лишена корковой пластинки, тогда как в базальной части ее выделяется разреженное скопление небольших полиморфных клеток (писleus septi).
Обонятельная луковица (bulbus olfactorius) может быть разделена на пять слоев: клубочковый (stratum glomerulosum), наружный волокнистый (stratum fibrosum externum), молекулярный (stratum moleculare), слой митральных клеток (stratum neurocitorum mitralium) и внутренний зернистый (stratum granulosum internum).
Миндалевидный комплекс, расположенный в вентральной части височной доли, может быть подразделен на два морфологически и функционально различных ядра – кортикомедиальное и базолатеральное. При цитоархитектоническом исследовании в нем отмечается наличие крупноклеточных, среднеклеточных и мелкоклеточных участков.
Амигдалярные клетки получают афференты от орбитальной части лобной доли, периформной коры, гипоталамуса, таламуса. Основными эфферентными системами амигдалы являются конечная полоска и вентральный амигдальный путь.
Эфферентные волокна через переднюю комиссуру достигают миндалевидного комплекса противоположной стороны. Другая часть волокон направляется в прозрачную перегородку. Кортикомедиальное и базолатеральное ядра взаимосвязаны между собой и отдают волокна в гипоталамус, к преоптической области и воронке. Некоторые аксоны клеток миндалевидного комплекса заканчиваются в хвостатом ядре.
Структуры миндалин играют существенную роль в интеграции инстинтивного пищевого и оборонительного поведения. Удаление или повреждение их вызывает нарушение эмоциональных реакций и неспособность к правильной оценке значения позитивных и негативных факторов внешней среды.
Нередко структуры старой и древней коры в целом относят к обонятельному ( или висцеральному) мозгу (rhinencephalon). Действительно, его передние компоненты, в том числе обонятельный бугорок и прямая извилина, являются первичными рецептивными зонами обонятельного анализатора в коре мозга. Кроме того, в него включают лимбические компоненты, не имеющие непосредственного отношения к восприятию запаховых раздражителей, хотя и тесно связанные с обонятельным анализатором и, вместе с тем, получающие афференты от других сенсорных систем. Это медиальная обонятельная область, серый покров мозолистого тела, а также гиппокамп. Лимбические структуры во взаимодействии с диэнцефалическими образованиями принимают участие в регуляции ряда вегетативных и соматосенсорных реакций, а также выполняют некоторые функции, необходимые для осуществления врожденных рефлексов и эмоционального поведения. Благодаря связям с ретикулярной формацией ствола обеспечивается комплексный характер нисходящих и восходящих влияний обонятельного мозга. В экспериментальных условиях его стимуляция оказывает тормозное влияние на стволовые механизмы, связанные с выражением эмоций (гнева, ярости и др.). Это происходит, в первую очередь, за счет подавления симпатических эффектов, исходящих из заднего гипоталамуса. С другой стороны, при повреждениях в передней лимбической и задней орбитальной областях возникают двигательное беспокойство, гиперактивность. Удаление передних частей гиппокампа и прилегающих участков коры, а также миндалевидного комплекса у животных приводит к появлению признаков гиперсексуальности и гиперфагии с утратой эмоциональных проявлений страха. Разрушение передних участков гиппокампа и миндалевидного комплекса, как показали наблюдения на кошках и обезьянах, сопровождается усилением реакций гнева и ярости.
Эти структуры принимают также участие в регуляции эмоциональных проявлений пищевого поведения.
Гиппокамп оказывает значительное влияние на гипифизарно-адреналовую систему.
Нейроны гиппокампа обладают особой чувствительностью в отношении многих инфекционных и токсических агентов. Дегенеративные изменения, приводящие к уменьшению в нем количества нейронов – частое явление при эпилепсии, нейросифилисе, отравлении окисью углерода, механических воздействиях.
Очевидно, гиппокамп получает часть обонятельных сигналов от нейронов прямой извилины и крючка через вставочные нейроны. Вместе с другими структурами старой коры он участвует своими волокнами в формировании таких проводящих путей, как свод, медуллярная и конечная полоски, а также медиальный пучок переднего мозга. В составе свода, наряду с частью аксонов гиппокампальных клеток, которые направляются в медиальную обонятельную область, содержатся также многочисленные волокна, идущие в противоположную сторону. Связи гиппокампа с субикулумом, пресубикулумом и энториальной корой также носят двухсторонний характер.
Кроме волокон, заканчивающихся в мамиллярных телах, гиппокамп образует связи и с другими гипоталамическими зонами, а также с некоторыми ядрами таламуса и покрышки среднего мозга.
Влияние гиппокампа и других структур старой и древней коры на функции неокортекса может обеспечиваться как благодаря прямым связям, так и опосредованно, через ретикулярную формацию ствола.
В целом, для лимбической системы характерно наличие замкнутых полисинаптических цепей. Например, одна из таких систем обеспечивает проведение от гиппокампа через мамиллярные ядра к переднему ядру таламуса, затем к коре поясной извилины и далее на субикулум, от которого импульсы вновь поступают в гиппокамп (рис. 7.13 и 7.15).
Благодаря таким круговым путям создаются предпосылки для циркуляции, необходимой для процессов запечатления информации.
У человека при некоторых психических заболеваниях, сопровождающихся нарушением кратковременной памяти, отмечаются дегенеративные изменения в гиппокампе. Так, характерным симптомом корсаковской амнезии является утрата способности запоминать только что происшедшие события, хотя давнее прошлое в памяти сохраняется.
Участие гиппокампа в механизмах кратковременной памяти и в процессах обучения обеспечивается благодаря обширной афферентации, поступающей от различных систем через зубчатую фасцию.
После удаления гиппокампа или перерезки зубчатой фасции особенно затрудняется выработка отставленных или задержанных условных рефлексов.
Поиск по сайту: