АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
Проводящие пути и связи таламических ядер
Таламические ядра средней линии получают волокна от других ядер таламуса и от ретикулярной формации ствола. Кроме того, они получают связи от основных соматосенсорных путей – спинноталамического и тригемино-таламического трактов, от мозжечка по пути его передних ножек и от четверохолмия по тектоталамическим путям. Эфферентные проекции обеспечиваются как внугриталамическими связями, так и волокнами от ядер средней линии к лимбической системе, включая структуры гипоталамуса и обонятельной коры. По-видимому, они передают сигналы также к полосатому телу в системе базальных ядер.
Среди ядер медиальной группы дорзальное ядро получает особенно многочисленные волокна от ядер средней линии. Кроме того, оно связано со всеми другими таламическими ядрами. Оно получает также афференты в виде части волокон медиальной петли от ядра тройничного нерва и от мозжечка. Дополнительно, в медиальном дорзальном ядре обнаруживаются волокна, выходящие из области перегородки и орбитальной коры лобной доли. Его основные эфферентные проекции прослеживаются в лобной коре, с которой оно, таким образом, имеет двусторонние связи. Это подтверждается тем, что после лобэктомии в нем наблюдается дегенерация, и его разрушение приводит к значительным изменениям в функции лобных долей. Обнаружено также, что часть эфферентов медиального дорзального ядра входят в состав гипоталамуса.
Меньше известно о связях второго большого ядра медиальной группы – центрального или срединного. Помимо обширных внутриталамических афферентов, оно получает входы от ретикулярной формации ствола, а также от проприоцептивных путей в составе спинальных и тригеминальных трактов. В свою очередь, это ядро отдает значительную часть эфферентов в полосатое тело. Некоторые исходящие из него волокна прослеживаются до среднего мозга.
Интраламинарные ядра медиальной группы, благодаря своим связям с ретикулярной формацией, включаются в нее и вместе с другими ретикулярными ядрами имеют обширное диффузное представительство в коре. В ядра дорзального отдела латеральной группы (дорзальное и срединное) направляется часть волокон от медиальной петли. Они получают также многочисленные внутриталамические связи – от вентральных и медиальных ядер. Основные эфференты их заканчиваются в теменной доле коры – в области задней центральной извилины (поля 1,2,3). Меньшая часть волокон заканчивается в области прецентральной извилины (поля 5,7). Описаны и обратные связи от теменной области коры к этим ядрам.
Ядра вентрального отдела латеральной группы, расположенные в задней части таламуса и объединяемые в вентро-базальный комплекс, являются основным переключательным звеном на пути тактильных, кинестетических и некоторых других сенсорных сигналов к коре мозга (рис. 7.29). Представительства нижних конечностей локализуются латерально, а от области головы – медиально. Вентральное заднелатеральное ядро этой группы является конечным путем медиальной петли и спинноталамического тракта. Концевые ветвления волокон медиальной петли, берущих начало от ядра тонкого канатика, распределяются назад и латерально по отношению к терминалям волокон клиновидного ядра. Окончания спинноталамического тракта в целом занимают более каудальную зону, чем концевые волокна медиальной петли.
Вентральное заднелатеральное ядро имеет связи с другими ядрами таламуса – центромедиальным, подушкой, а также с субталамическим ядром. Основным местом окончания аксонов этого ядра является область задней центральной извилины в коре мозга (1,2 и 3 поля). Кроме того, часть эфферентных волокон от него направляется к полосатому телу и к субталамусу. Вентральное постеромедиальное (полулунное) ядро получает волокна тройничного нерва, идущие в составе вентрального тригеминоталамического тракта (тригеминальной петли). Как и постеролатеральное ядро, оно проецируется на кору с сохранением топографического представительства поверхности тела. Эфферентные волокна от него проходят через заднюю дугу внутренней капсулы и распределяются по корковым зонам задней центральной извилины (поля 1,2,3). Дополнительно, оно имеет внутриталамические связи с центромедиальным ядром и подушкой. Соседнее с ним добавочное полулунное ядро является местом окончания вторичных вкусовых волокон. Оно также имеет топическое представительство в коре больших полушарий (в 43 поле) (рис. 7.10; 7.14; 7.16 и 7.31).
Переднее вентральное ядро латеральной группы получает волокна от одного из ядер полосатого тела – бледного шара – через посредство таламического пучка и чечевичной петли. По-видимому, часть афферентов этого ядра берет начало в зубчатом ядре мозжечка. Оно отдает эфферентные волокна к полосатому телу. Некоторые волокна проходят дальше – в переднюю центральную извилину коры (поле 6) и, по-видимому, в кору островковой доли. Взаимодействие с другими ядрами промежуточного мозга обеспечивается за счет его связей с медиальной группой и субталамическим ядром.
Латеральное вентральное промежуточное ядро получает большую часть афферентных волокон от зубчатого ядра противоположной стороны, восходящих в составе верхних ножек мозжечка. В него заходят, кроме того, некоторые волокна таламического пучка, берущие начало от бледного шара и частично волокна от красного ядра (рис. 7.31).
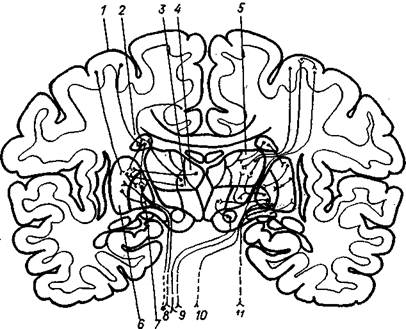
Рис. 7.31. Афферентные и эфферентные связи базальных ядер (слева – палеостриатума, справа – неостриатума):
1 – кора большого шара; 2 – хвостатое ядро; 3 – итраламинарные таламические ядра; 4 – медиальные таламические ядра; 5 – латеральные, вентральные и передние ядра таламуса; 6 – скорлупа; 7 – бледный шар; 8 – связи с черной субстанцией; 9 – волокна к красному ядру; 10 – волокна к нижней оливе и ретикулярной формации среднего мозга; 11 – связи с полем Н и субталамическим ядром
Эфференты этого ядра направляются через заднюю дугу внутренней капсулы к коре передней центральной извилины (4 и 6 поля) (рис. 7.10 и 7.14).
В наружном коленчатом теле оканчивается большая часть перекрещенных и неперекрещенных волокон зрительного тракта, поворачивающих сбоку у верхнего конца мозговых ножек. Они распределяются по различным слоям дорзальной части коленчатого тела в соответствии с картиной ретинотопической проекции. Кроме того, наружное коленчатое тело получает афференты из среднего мозга через ручки верхних бугорков. В вентральную часть его входят волокна от дорзальной части. Волокна, выходящие из слоистой дорзальной части этого ядра, восходят вначале в составе задней дуги (ретролентиформной части) внутренней капсулы и затем зрительной радиации (radiatio optica) на медиальную поверхность затылочной доли коры, заканчиваясь в области шпорной борозды (s. calcarinus). Меньшая часть эфферентов наружного коленчатого тела направляется вниз в верхние бугорки и в неопределенную субстанцию субталамуса. Кроме того, наружное коленчатое тело имеет связи с другими таламическими ядрами – подушкой, дорзальным и вентральным отделами латеральной группы.
Ядро внутреннего коленчатого тела служит местом окончания волокон латеральной петли. Таким образом, оно получает сигналы от улиткового ядра, ядра латеральной петли, трапециевидного ядра и верхней оливы. Кроме того, в него приходят волокна, начинающиеся в нижнем двухолмии и идущие в составе его ручек.
Аксоны клеток внутреннего коленчатого тела направляются и заканчиваются в области передней поперечной извилины височной доли (41 поле). При этом на всем пути сохраняются тонотопичсские проекционные отношения (рис. 7.14; 7.16 и 7.17).
Значительно менее многочисленны исходящие из внутреннего коленчатого тела волокна, которые достигают субталамуса, а также гомо- и контралатсрального задних бугорков крыши среднего мозга.
Между собой коленчатые тела обеих половин таламуса имеют межъядерные связи.
Ядро подушки включено в систему кольцевых связей с передним мозгом, корой больших полушарий, а также с соседними ядрами таламуса. В первую очередь, это волокна, которые оно посылает в теменную, височную и затылочную кору (поля 22, 42, 18 и 19) и направляющиеся к нему афференты от этих долей больших полушарий. Сюда относятся, кроме того, связи с коленчатыми телами, ядрами латеральной группы таламуса и с четверохолмием.
Передняя ядерная группа имеет межъядерные связи с медиальными и латеральными ядрами своей стороны и внутриталамические связи с противоположной стороной через комиссуральные волокна. Таким образом, они могут получать сигналы соматосенсорной и кинестетической чувствительности, переключаемые в вентролатеральных ядрах. В этой группе заканчиваются гипоталамические волокна, идущие в составе мамилло-таламического пучка. Связи одного из ядер этой группы – переднего дорзального с мамиллярными ядрами, видимо, носят двухсторонний характер. От главного, переднего, или вентрального ядер берут начало эфферентные пути к поясной извилине (gyrus cinguli) коры (поля 25 и 24) (рис. 7.13; 7.14 и 7.30).
Поиск по сайту: