АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
Оболочка клетки
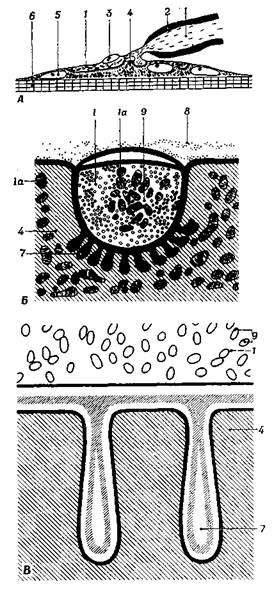
Рис. 2.3. Нервно-мышечный синапс:
А – схема концевой двигательной пластинки. Концевые веточки нервного волокна лежат в "синаптических ямках ", и обе структуры образуют нервно-мышечный синапс. Непосредственно под границей раздела аксоплазмы и саркоплазмы находятся лентовидные субневральные пластинки, которые в поперечном сечении имеют форму палочек; Б – синаптическая "ямка", или "желобок", на поперечном разрезе; В – две складки в области нервно-мышечного соединения; видно, как поверхностная саркоплазматическая мембрана соединяется с поверхностной аксоплазматической мембраной, образуя сложную синаптическую мембрану; 1 – аксоплазма (с митохондриями); 1a – митохондрия; 2 – миелиновая оболочка; 3 – шванновская клетка в области нервно-мышечного соединения; 4 – саркоплазма (с митохондриями); 5 – ядро мышечной клетки; 6 – миофибриллы; 7 – складка нервно-мышечного соединения; 8 – коллагеновые волокна; 9 – пузырьки
Рис 2.3. До появления электронной микроскопии о структуре клеточной оболочки было известно очень мало. Теперь мы знаем, что она представляет собой непрерывную сдвоенную мембрану, окружающую целиком тело клетки, дендриты и аксон. Помимо этого, аксон дополнительно окружен серией сложных липопротеидных пластин, спирально обернутых вокруг него, которые в периферической нервной системе образованы шванновскими клетками, а в центральной – клетками олигодендроглии. Клеточная оболочка состоит из двух электроноплотных слоев, разделенных бесструктурной промежуточной зоной. Следовательно, в определенном смысле это трехслойная оболочка, общая толщина которой равна примерно 90– 100 А. Оказалось, что эта оболочка геометрически асимметрична и что на ее внутренней, цитоплазматической стороне расположен более толстый и темный электроноплотный слой, а на наружной – более тонкий. Подобные же сдвоенные электроноплотные мембраны можно видеть и в клетке. Ядерная оболочка, эндоплазматическая сеть, комплекс Гольджи и митохондрии также образованы двойными мембранами. Мембраны комплекса Гольджи и митохондрии более симметричны, чем наружная плазматическая мембрана, и часто содержат окрашивающееся вещество в среднем "светлом" слое. Окружающие аксон мембраны шванновских клеток, мезаксон, связанный с миелиновой оболочкой, и синаптические мембраны также двухслойные (или трехслойные, если принимать светлую промежуточную зону за истинный слой). Термин "мезаксон" относится к тому отделу, где края цитоплазматической мембраны шванновской клетки смыкаются вокруг заключенного в них аксона и образуют дубликатуру. Аксон в известном смысле подвешен в складке цитоплазматической мембраны наподобие того, как в брюшной полости позвоночных кишка подвешена на дубликатуре брюшины – брыжейке (mesenterium) <...>. В области концевой двигательной пластинки, где плазматическая мембрана нейрона находится между аксоплазмой нервного волокна и саркоплазмой мышечного, мембрана эта несколько видоизменена. Здесь центральный участок между оболочками занят плотным веществом, увеличивающим общую толщину аксосаркоплазматической мембранной структуры до 500–700 А. Робертсон <...> обозначает это образование как синаптический мембранный комплекс, поскольку точно не ясно, какие слои следует обозначить в данном случае термином "клеточная оболочка".
На раннем этапе развития электронной микроскопии была выдвинута концепция "элементарной мембраны". Эта концепция исходила из допущения, что все клеточные мембраны имеют единый план строения, и на этом основании все внутриклеточные мембранные структуры можно было представить происходящими из наружной клеточной мембраны, как это показано на рис. 2.4. Эта концепция была основана на представлении о том, что все двухслойные мембраны имеют равную толщину и симметрию. Поскольку многочисленные исследования выявили различия в строении мембран митохондрий, аппарата Гольджи и a-цитомембран <...>, в настоящее время гипотеза "элементарной мембраны" представляет, по-видимому, лишь исторический интерес. Несомненно, что отказ от этой гипотезы должен повлечь за собой дальнейшие исследования, призванные выяснить вопрос об источниках и процессах формирования внутриклеточных мембранных структур. Хотя происхождение таких мембранных систем неясно, наличие каналов эндоплазматической сети, которые, как полагают, сообщаются с внеклеточным пространством, общепризнано. Можно полагать, что эти каналы представляют "циркуляторную систему" клетки, которая облегчает поступление в нее питательных веществ и выход секретируемых продуктов и конечных продуктов обмена. Есть также наблюдения, показывающие, что эндоплазматическая сеть соединена с наружным слоем ядерной мембраны. Помимо этого, отверстия, или поры, в ядерной мембране служат тем путем, по которому идет обмен между нуклеоплазмой и цитоплазмой. Комбинация эндоллазматической сети и рибосом представлена в нейронах веществом Ниссля и является эквивалентом гранулярной эндоплазматической сети, описанной в других клетках. Кроме рибосом, связанных с эндоплазматической сетью, для нейронов характерно также и огромное число свободных рибосом, которые часто образуют скопления, или "розетки". Они особенно многочисленны в дендритах и перикарионе.
Ядро
Рис. 2.4. Ядро (1) нейрона имеет большие размеры и по сравнению с ядрами других клеток окрашивается слабо. Поскольку количество ДНК в нем такое же, как и в ядрах клеток печени, селезенки, мышц и поджелудочной железы (2,3 × 10-2 г), слабую базофилию можно объяснить дисперсией ДНК в относительно большом объеме. Ни световой, ни электронный микроскопы не выявляют в ядрах зрелых нейронов никаких признаков присутствия хромосом.
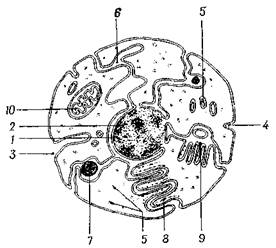
Рис. 2.4. Схема, иллюстрирующая концепцию Робертсона, согласно которой все цитоплазматические органеллы образуются из наружной клеточной мембраны:
1 – ядро; 2 – ядрышко; 3 и 4 – ямка; 5 – эндоплазматическая сеть; 6 – внутриклеточное пространство нейрона, сообщающееся с внеклеточным пространством; 7 – секреторная гранула; 8 – рибосомы; 9 – аппарат Гольджи с капельками секрета; 10 – митохондрия
Электронная микроскопия выявляет в двухслойной ядерной мембране множество отверстий, или пор, которые, по-видимому, обеспечивают обмен между нуклеоплазмой и цитоплазмой. Возможно, эти поры служат для выхода новообразованной РНК из ядра в цитоплазму, где она включается в состав вещества Ниссля (5).
В ядре нейрона обычно находится ядрышко (2), которое характеризуется высокой степенью базофилии. Ядрышко содержит большое количество РНК и покрыто тонким слоем ДНК. У человека ядрышко интенсивно окрашивается люксолевым синим прочным, который выявляет холинсодержащие липиды. Многочисленные исследования выявили связь ядрышка с синтезом рибонуклеопротеидов в те периоды, когда в нейроне осуществляется синтез белка. Значение реакции с люксолевым синим прочным пока не выяснено.
Хиден <...> показал, что ядрышко играет ведущую роль в жизни клетки, особенно в синтезе нуклеиновых кислот и белков. Наблюдения Тьюэри и Бурне <... > также показывают, что ядрышки клеток спинномозговых узлов (у крысы) являются центрами высокой метаболической активности и, возможно, участвуют в синтезе разнообразных ферментных белков. С помощью гистохимических реакций эти авторы обнаружили в ядрышке АТФазу, глюкозо-6-фосфатазу, сукцинатдегидрогеназу, щелочную фосфатазу, глюкозофосфорилазу, 5-нуклеотидазу и специфическую холинэстеразу <...>. Интенсивность этих гистохимических реакций, различная у разных клеток, по-видимому, зависит от активности ядрышка. Подобное обилие ферментов связано с дыхательной, энергетической и синтетической функциями нейрона. Конечно, выявляются и другие ферменты, но именно эти указывают на связь ядрышка с функциональными проявлениями нейронов, активное участие в которых принимают также аппарат Гольджи (9) и митохондрии (10).
Известно также, что ядрышки бывают разного размера – от небольших телец до крупных сложных образований, почти целиком заполняющих ядро. Электронно-микроскопические исследования позволяют предположить, что эти различия в размере ядрышка связаны с физиологическим состоянием клетки <...>. В то же время многие исследования показали, что размер ядрышка биохимически связан с его синтетической активностью. Наблюдения Тьюэри свидетельствуют о том, что существует связь между синтезом РНК, белков и липидов в ядрышке и переносом этих веществ в цитоплазму нервной клетки в тот момент, когда ядрышко оказывается вблизи ядерной мембраны. Эти наблюдения подтверждают более ранние данные Эйнарсона <...>, свидетельствующие о том, что вещество Ниссля (5) в нейроне исходит из ядрышка.
Структурно ядрышко представляет собой плотное внутриядерное скопление, состоящее из РНК, белков и липидов, а также некоторого количества ДНК. Является ли оно дискретной структурой или нет – вопрос спорный, поскольку у ядрышка нет выраженной пограничной мембраны. Постоянные изменения его размеров и положения, его гистохимических реакций, связанные с изменениями физиологического состояния нейрона, свидетельствуют об исключительно высокой пластичности этой органеллы и ее важном значении для синтеза РНК и различных ферментов.
В ряде электронно-микроскопических работ были описаны фибриллярные внутриядерные включения, имеющие решетчатую структуру и форму дисковидной пластинки диаметром 1,8 мкм и толщиной 0,1 мкм. Функция этих включений неизвестна.
Поиск по сайту: