АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
СПАДКОВІСТЬ
|
Читайте также: |
Згадаєте! Що таке ген, геном, генотип, генетичний код, рекомбінація?
Поняття про ген. Раніше ви довідалися про хімічну природу гена. Термін «ген» був запропонований в 1909 році датським ученим В. Ио-ганнсеном (1857-1927). Тривалий час, поки не були з'ясовані структура нуклеиновых кислот і генетичний код, ген уважали неподільною одиницею спадкоємної інформації, рекомбінацій і мутацій. Але згодом установили, що мутації можуть торкати не всього гена, а тільки певну його частину; у вре-
мя кроссинговера гомологичные хромосоми можуть обмінюватися як окремими генами, так і їхніми частинами. Мінімальна ділянка молекули нуклеиновой кислоти, що може бути розділений під час кроссинговера, становить усього 1-2 нуклеотидные пари. Однак ген - цілісна одиниця, оскільки будь-які порушення його структури змінюють закодовану в ньому інформацію або приводять до її втрати.
Серед генів виділяють структурні, що кодують структуру білків і рибонуклеиновых кислот, і регуляторні, що служать місцем приєднання ферментів і інших біологічно активних з'єднань, що впливають на активність структурних генів і приймаючу участь у процесах подвоєння ДНК і транскрипції. Розміри регуляторних генів, у порівнянні зі структурними, як правило, незначні.
Таким чином, ген - спадкоємний фактор, функціонально неподільна одиниця генетичного матеріалу у вигляді ділянки молекули нуклеиновой кислоти (ДНК або РНК), що кодує первинну структуру поліпептиду, молекули транспортної або рибосомальной РНК, або взаємодіючого з біологічно активними речовинами (напр., ферментами).
Організація генома в різних організмів. У різних організмів кількість генів у геноме може варіювати. Найбільше просто організований геном вірусів, що може включати від одного до декількох сотень генів. Геном прокаріот улаштований більш складно й містить як структурні, так і регуляторні гени. Наприклад, ДНК кишкової палички складається з 3 800 000 пар нуклеотидов, а кількість структурних генів наближається до однієї тисячі. Майже половина довжини молекули ДНК генетичної інформації не несе, це ділянки, що лежать між генами.
Геном эукариот має ще більш складна будова: у їхніх ядрах
:ольше втримується ДНК, отже більше й кількість
гтруктурных і регуляторних генів. Так, геном дрозофилы з-
:тоит майже з 180 000 000 пар нуклеотидов і нараховує око-
: до 10 000 структурних генів; у геноме людини - понад 100 000
I структурних генів.
Дослідження генома різних організмів показали, що ко-|дгчество ДНК у ядрі в 8-10 разів перевищує необхідне для ко-щкрования всіх структурних генів. Причини цього явища раз-■шг-шые. По-перше, ДНК эукариот містить велика кількість ■сследовательностей нуклеотидов, кожна з яких повторя-■гя до сотень тисяч разів. По-друге, значна частина ДНК в-■§~ie не несе генетичної інформації. По-третє, є рг-: жество регуляторних генів, що не кодують структуру поли-Ьегтидов або РНК.
-• -астки що не кодує ДНК виявлені й у складі структурних генів. Було Казано, що гени складаються з окремих блоків, одні йз яких (экзоны) ■имоуются в иРНК і несуть інформацію про структуру певних соедине-
ний, а інші (интроны) - немає. Окремі интроны можуть включати від 100 до 1 000 000 і більше нуклеотидных пара. Кількість интронов усередині окремих генів може бути різним: у гені, що кодує структуру гемоглобіну, їх 2, яєчного білка - 7, белка-коллагена курки - 51. Кількість і порядок розташування интронов специфічні для кожного гена. Гени копіюються в молекулі-попередниці ирнк (про-ирнк), звідки интроны віддаляються за допомогою особливих ферментів, а экзоны залишаються й з'єднуються в строго певному порядку. Так утвориться зріла ирнк.
Цитоплазматичёская спадковість. У клітинах эукариот, крім спадкоємного матеріалу, розташованого в ядрі, виявлена також внеядерная, або цитоплазматическая спадковість. Вона зв'язана зі здатністю певних структур цитоплазми зберігати й передавати нащадкам частина спадкоємної інформації батьків. Хоча провідна роль у спадкуванні більшості ознак організму належить ядерним генам, цитоплазматическая спадковість значно впливає на прояв деяких спадкоємних ознак. Вона пов'язана із двома видами генетичних явищ:
- спадкуванням ознак, кодируемых внеядерными генами, розташованими в певних органеллах (мітохондріях, пластидах);
- проявом у нащадків ознак, обумовлених ядерними генами батьків, але на формування яких впливає цитоплазма яйцеклітини.
Існування генів в органеллах (мітохондріях і пластидах), здатних до подвоєння, установлено ще на початку XX ст. при вивченні зелених і безбарвних пластид у деяких квіткових рослин, що мають мозаїчне фарбування листів. Внеядерные гени взаємодіють із ядерними й перебувають під контролем ядерної ДНК. Цитоплазматическая спадковість, пов'язана з генами пластид, відома в таких рослин, як левиний зев. Серед цих рослин є форми зі строкатими листами, причому ця ознака передається по материнській лінії. Строкатість листів обумовлена тим, що частина пластид не здатна утворювати хлорофіл. Після розподілу клітин з безбарвними пластидами в листах виникають білі плями, що чергуються із зеленими ділянками. Передача пестролистости по материнській лінії пояснюється тим, що під час утворення полових клітин пластиди попадають у яйцеклітини, а не в спермін. Пластиди, що розмножуються розподілом, мають генетичну безперервність: зелені пластиди дають початок зеленим, а безбарвні - безбарвним. Під час розподілу клітини пластиди розподіляються випадково, у результаті чого утворяться клітини з безбарвними, зеленими або обома типами пластид.
Явище цитоплазматической спадковості, пов'язане з генами мітохондрій, вивчали на прикладі дріжджів. У цих мікроорганізмів у мітохондріях виявлені гени, що спричиняються відсутність або наявність дихальних ферментів, а також стійкість до певних антибіотиків.
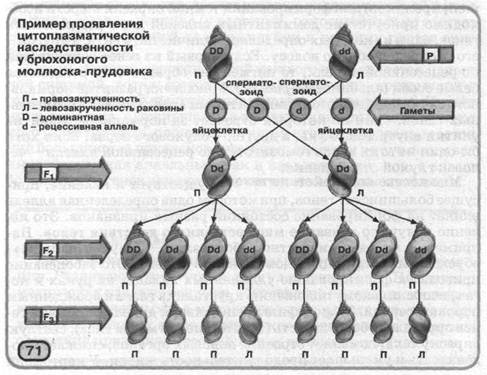
Вплив ядерних генів материнського організму через цитоплазму яйцеклітини на формування певних станів ознак можна простежити на прикладі прісноводного брюхо-ногого молюска - прудовика {мал. 71). У нього є форми з різними станами спадкоємної ознаки - лівого або правого напрямку закрученности раковини. Аллель, що визначає правозакрученность раковини, домінує над аллелью левозакрученности, але при цьому напрямок закрученности визначається генами материнського організму. Наприклад, особини, гомозиготні по рецессивной аллели левозакрученности, можуть мати правозакрученную раковину, якщо вони відбулися від материнського організму, що мав домінантну аллель правозак-рученности.
Відносини «ген — ознака». Тривалий час у генетику існувало правило, відповідно до якого кожний ген визначає синтез одного поліпептиду («один ген - одна ознака»). Однак подальші дослідження показали, що відносини «ген - ознака» значно складніше, ніж уявляли собі раніше. Стали відомі явища множинної дії генів і взаємодії неаллельных генів.
Раніше ми розглядали випадки взаємодії аллельных генів: повне й неповне домінування, проміжний характер спадкування. Однак часто на формування певних з-
стоянь ознак впливає взаємодія двох або більше неал-лельных генів, що може здійснюватися в різних формах.
Один з типів взаємодії неаллельных генів проявляється в тім, що певна аллель одного гена придушує прояв аллели іншого, неаллельного. Такий тип взаємодії неаллельных генів називається придушення. Наприклад, пурпурне фарбування очей дрозофилы визначаються рецессивной аллелью. Але вона не виявиться у фенотипі, якщо в гомозиготних по цієї аллели особин також у гомозиготному стані є присутнім рецессивная аллель іншого неаллельного гена, що придушує прояв першої.
Інший тип взаємодії неаллельных генів - це доповнення, або комплементарностъ. Для нього необхідна взаємодія домінантних аллелей двох або більше неаллельных генів, які разом визначають прояв конкретного стану ознаки. Так, фіолетове фарбування плодів баклажана залежить від взаємодії домінантних аллелей двох неаллельных генів, завдяки якому утвориться відповідний пігмент. Якщо хоча б один із цих генів гомозиготен по рецессивной аллели, пігмент не синтезується й плоди будуть безбарвними.
Подібне спостерігається й у запашного горошку, у якого домінантні аллели двох неаллельных генів спричиняються червоне фарбування віночка (якщо хоча б один з генів гомозиготен по рецессивной аллели - віночок білий). У тварин (напр., мишей, кроликів) для формування темного фарбування вовни необхідна присутність домінантних аллелей двох неаллельных генів, один із яких визначає наявність пігменту, а іншої -його розподіл по волоссю. Якщо перший з генів гомозиготен по рецессивной аллели, то пігмент не утвориться й народжуються білі особини (альбіноси) {мал. 72). У людини розвиток нормального слуху визначається домінантними аллелями двох неаллельных генів, один із яких відповідає за нормальний розвиток равлика внутрішнього вуха, а іншої - слухового нерва. Якщо хоча б один із цих генів гомозиготен по рецессивной аллели - людина глухий від народження.
Множинна дія генів. Існує і явище, властивій більшості генів, при якому одна певна аллель впливає на формування станів різних ознак. Це явище одержало назву множинної дії генів. Наприклад, у людини відоме захворювання - «павукові пальці», обумовлене мутацією домінантної аллели. Це захворювання приводить до формування подовжених пальців на руках і ногах, неправильному положенню кришталика ока й уроджених пороків серця. У дрозофилы рецессивная аллель одного з генів визначає белоглазость (відсутність пігменту око), світле фарбування тіла, змінює будову полових органів, знижує плідність і зменшує тривалість життя. У картоплі
• " ""■"■""""-—-~~—■• "*""*и-™и""™||™|в1™1*————»——«.
Приклад Взаємодії /4# сірий В1^ білий
неаллельных. генів. >*Чр^яИ1^ь. J/lJs^*"***^
у кроликів СЩ^г Щк (Шгг s~* Л
~]рГ > ССАА Х\ ссаа
-:-'---------- v |_____I I
/^И1 сірі
Jmj ^сірі //^7 білі ^Lr чорні
ССАА I...........| ^ряязн ^^^
|=MF I--------~\ ССАа ****! &
I I 21______,/ СсАА 9/16 ссАа 4/16 ССаа я/16
^Т I СсАа ссаа Ссаа
А - аллель, що визначає концентрацію пігменту в підстави волоска; ^ v a - його рівномірний розподіл
Г 72 J ^ ~ аллель, що визначає наявність пігменту;
Поиск по сайту: