АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
БІОГЕОГРАФІЯ Й ЕВОЛЮЦІЯ
Згадаєте! Що таке ареал? У чому сутність понять «географічна й екологічна ізоляція»? Що собою представляють підвиди?
Біогеографічне районування біосфери Біогеографія -
наука про закономірності поширення на земній кулі видів і інших систематичних груп живих організмів і їхніх екологічних угруповань.
Вивчаючи ендемічні види, пологи й ін. систематичні групи, а також основні типи біогеоценозів, учені виявили, що різні частини біосфери відрізняються між собою.
Эндемики (від греч. эндемос - місцевий) - систематичні групи (вид, рід, сімейство й ін.), що живуть тільки в певній частині біосфери. Наприклад, бізон - эндемик Північної Америки, сосна кримська - Криму.
Хоча зоологічне й ботанічне районування близькі між собою, однак не повністю збігаються. Це обумовлено здатністю тварин до активних міграцій і значним впливом особливостей ґрунту, клімату й ін. факторів на рослини, ведучих прикріплений спосіб життя.
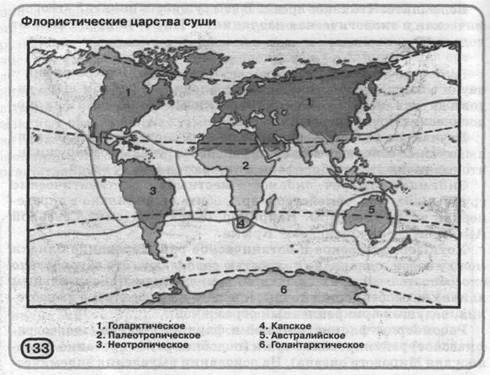
Розглянемо флористичне й фауністичне (зоогеогра-фическое) районування суши (подібне районування створене й для Світового океану). На підставі виділення эндемичес-
ких комплексів ботаніки ділять сушу на б царств (мал. 133), у кожному з яких виділено кілька областей (усього їх 34), а в останніх - близько 150 провінцій. Україна перебуває в межах Циркумбореальной області Голарктического царства. Сучасне флористичне районування Землі розробив відомий вірменський учений А. Л. Тахтаджян. Зоогеографы виділяють 7 фауністичних областей (мал. 134).
Значення біогеографії для розвитку еволюційних поглядів. Дані біогеографії використають для обґрунтування положення про географічну ізоляцію як необхідної умови видоутворення (згадаємо про дослідження Ч.Дарвіна розмаїтості вьюрков на островах Галапагос). Однак зараз зібраний великий фактичний матеріал, що свідчить про те, що ізоляція далеко не завжди викликає дивергенцію й видоутворення.
Більшість видів прісноводних і ґрунтових одноклітинних тварин поширено по усьому світі; ті самі види ногохвосток (дрібні безкрилі ґрунтові членистоногие) живуть на більшості континентів. Отже, незважаючи на нездатність до активних міграцій або поширення іншими способами, популяції таких видів, тривалий час перебуваючи в повній ізоляції, не дивергируют і не перетворюються в нові види. Це свідчення того, що екологічна ніша виду зберігає його цілісність у стабільних умовах існування

незалежно від можливості обміну спадкоємною інформацією між популяціями, а також форм розмноження.
Дані біогеографії підтверджують єдність походження флори й фауни певних царств, областей і ін. ендемічних комплексів. Наприклад, у різних областях Голарктического царства існують подібні біогеоценози різних кліматичних зон (від тундри до субтропіків), до складу яких входять близькі види (звичайний і американський клени, європейські й американські норка й бобер і ін.). Ці факти вказують на існування в порівняно недалекому минулому сухопутних зв'язків між Північною Америкою і Євразією. Дійсно, геологи довели, що протягом різних періодів і епох кайнозойской ери існували такі «мости» через Гренландію й Ісландію з Європою й через сучасний Берингове протоку - з Азією.
Аналіз числа видів тієї або іншої систематичної групи в межах різних біогеографічних регіонів дозволяє встановити центр її видової розмаїтості (територія з найбільшим числом видів). Так, центром видової розмаїтості сумчастих ссавців є Австралія, кактусів - Центральна й Південна Америка. Раніше вважали, що центри сучасної видової розмаїтості одночасно є й місцями історичного походження систематичних груп, але дані палеонтології й геології зруйнували це подання.
Наприклад, сучасні мозоленогие (загін ссавців) у дикому стані живуть лише в пустелях Центральної Азії (двогорбий верблюд) і горах Південної Америки (лами). Однак на підставі палеонтологічних даних було встановлено, що цей загін виник у Північній Америці, звідки його види через сухопутні «мости» розселилися в Південну Америку й Азію. У Північній Америці вони вимерли через практично повне заледеніння її території в антропогенний період.
Дані біогеографії, палеонтології й геології дозволили переосмислити традиційні подання про причини вимирання одних груп організмів і широкий адаптивной радіації видів, що зайняла їхні місця.
Дарвіністи затверджують, що так звані примітивні групи витісняються більше високоорганізованими «прогресивними». У сутності це - подальший розвиток навчання Ж.-Б.Ламарка про градації. Тим часом підвищення рівня організації, як способу досягнення біологічного прогресу, спостерігається тільки в деяких філогенетичних галузей (насінні рослини, молюски, членистоногие, хордові), причому усередині кожної з них процвітають різні за рівнем складності групи (напр., всі класи хребетних, починаючи з «примітивних» хрящових риб і земноводних). Такі дуже розповсюджені й богатые видами типи тварин, як кишечнополостные, круглі й кільчасті хробаки; відділи рослин - червоні й бурі водорості, мохоподібні та ін. із часу свого виникнення ніяк не ускладнилися; а біологічний прогрес у величезного числа паразитичних видів (включаючи цілі типи й класи) був досягнутий завдяки загальній дегенерації.
Ч.Дарвін уважав, що сумчасті ссавці були спочатку поширені на всій суші, але згодом були витиснуті більше «прогресивними» плацентарними й в Австралії збереглися лише завдяки її тривалій ізоляції від інших континентів. Дослідження американського вченого Л.Маршалла (1980) повністю спростували це твердження. Як свідчать дані палеонтології, сумчасті виникли в Америці близько 100 млн років тому. Пізніше Північна й Південна Америка втратили сухопутний зв'язок, і еволюція сумчастих у цих двох частинах миру відбувалася різними шляхами. На початку кайнозойской ери сумчасті розселилися з Північної Америки в Європу через існуючий тоді сухопутний «міст». Вони тривалий час співіснували там із плацентарними й бурхливо еволюціонували на цих континентах аж до антропогенного періоду, коли вимерли внаслідок заледеніння Північної півкулі.
Приблизно 12 млн років тому значне заледеніння відбулося в Південній півкулі, що привело, зокрема, до майже повного вимирання флори й фауни Австралії. Трохи раніше через сухопутні «мости» між Південною Америкою, Антарктидою (тоді вона мала досить теплий клімат) і Австралією в останню мігрували сумчасті. заледеніння, Що Пішло, вони пережили в її північних областях і потім заселили весь материк внаслідок широкої адаптивної радіації. Плацентарні не змогли заселити цей континент, тому що всі сухопутні адаптаційні зони ссавців уже були зайняті сумчастими.
Отже, основною причиною вимирання видів є розпад біогеоценозів, до складу яких вони входили. Зрозуміло, що насамперед вимирають високоспеціалізовані види з низькою екологічної (еволюційної) пластичністю.
При освоєнні нових ділянок (напр., новостворених островів) або місць зруйнованих экосистем важливе значення має послідовність вселення. Перші види (піонери), що проникнули на такі ділянки, внаслідок достатку життєвих ресурсів і відсутності конкуренції з екологічно близькими видами повністю освоюють певну адаптаційну зону, образуя максимально можливе число екологічних ніш. Наприклад, результати дослідження фауни комах геологічно дуже молодих (виникли біля мільйона років тому) Гавайських островів показали, що перші види (піонери) дали величезну кількість ендемічних видів (один рід вусань^-вусанів-жуків-вусанів - 136, мухи-дрозофилы - більше 300).
Поиск по сайту: