АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
ПОПУЛЯЦІЯ ЯК ЕЛЕМЕНТАРНА ОДИНИЦЯ ЕВОЛЮЦІЇ
Згадаєте! Які основні положення генетики популяцій?
Особливості взаємодії генотипів і фенотипів у популяціях. Як відомо, різні фактори навколишнього середовища впливають на фенотип, оскільки генотип - це лише закодована в ДНК спадкоємна інформація. Остання часто не проявляється у фенотипі, якщо перебуває в рецессивной аллели в гетерозиготному стані. Тому в популяціях перехресно запліднюються диплоидных організмів рецессивные мутації тривалий час можуть не проявлятися, перебуваючи в гетерозиготному стані й образуя резерв спадкоємної мінливості.
Ніж ширше такі мутації поширюються серед особин популяції внаслідок послідовних схрещувань, тим більше ймовірний їхній перехід у гомозиготний стан і прояв у фенотипі. Залежно від відповідності таких станів ознак певним умовам середовища їхні носії - організми -або гинуть, або виживають і дають плідне потомство. В останньому випадку через кілька поколінь такий стан ознаки (фен) як більше пристосувальне повністю витисне з популяції менш відповідний даним умовам існування домінантний фен разом з його носіями (особинами). Таким чином, внаслідок подібних еволюційних змін зберігається тільки рецессивная мутантна аллель, а її домінантна аллель зникає.
Напівдомінантні й домінантні мутації внаслідок взаємодії організму - їхнього носія - з навколишнім середовищем у процесі онтогенезу в тім або іншому ступені проявляються у фенотипі. Якщо даний фен не відповідає умовам навколишнього середовища, то ці особини або відразу елімінують (гинуть) або не

дають плідного потомства; якщо він надає організму певні переваги, те такий фен широко поширюється в популяції в процесі зміни поколінь.
Таким чином, незважаючи на природу (рецесивність або домінантність) того або іншого фена, він при відповідності умовам навколишнього середовища поширюється в популяції внаслідок розмноження своїх носіїв - особин, або, навпаки, зникає разом з фенотипами, до яких він належить. Одночасно здійснюється й перерозподіл частот зустрічальності
окремих генів.
Спостереження в природі підтвердили здатність популяцій пристосовуватися до змін середовища перебування. Так, у середині XIX ст. в Англії переважали особини метелика березового п'ядака світлого фарбування, що відповідає тлу стовбурів беріз, на яких вони сидять удень. У промислових районах до кінця сторіччя внаслідок задимленості стовбури беріз стали сірими, і через кілька десятиліть у популяціях березового п'ядака початку домінувати темноокрашенная форма {мал. 124). Останнім часом внаслідок боротьби із промисловими викидами в атмосферу стовбури беріз в Англії знову стали білими, а серед метеликів знову переважають светлоокрашенные особини, тому що в їхніх популяціях стала більше адаптивної до даних умов аллель світлого фарбування.
Популяція — елементарна одиниця еволюції. Синтетична гіпотеза розглядає популяцію як елементарну одиницю еволюції, тому що в ній відбуваються всі елементарні еволюційні процеси. Окремі особини популяції не мають своєї власної еволюційної долі, тому що всі еволюційні події відбуваються під час зміни поколінь. Відповідно до синтетичної гіпотези, у популяціях, крім спадкоємної мінливості, добору й боротьби за існування, діють ще й особливі елементарні фактори еволюції - дрейф генів, хвилі життя й ізоляція. Про дрейф генів вам уже відомо з § 48 дійсного підручника, два інших поняття розглянемо докладно.
Хвилі життя - це коливання чисельності особин популяції. Вони можуть залежати від особливостей життєвого циклу виду й визначатися генетично, наприклад, сезонні зміни чисельності комах, однолітніх рослин. Крім того, хвилі життя можуть бути викликані змінами інтенсивності дії екологічних факторів (умови зимівлі, тиск хижаків і паразитів, наявність кормових ресурсів та ін.). Зміни чисельності можуть бути періодичними або відбуватися без видимого порядку.
Еволюційне значення хвиль життя полягає в тім, що зміна співвідношень певних аллелей у генофонді при коливаннях чисельності особин не залежить від ступеня пристосованості організмів. Наприклад, у сувору зиму вимерзають організми верхніх шарів ґрунту й виживають лише ті, хто випадково виявився в місцях з менш інтенсивним промерзанням (у місцях, де вітер не здуває сніг і т.п.). Хвилі життя - одна із причин дрейфу генів.
Ізоляція - це фактор, що утрудняє або робить неможливим схрещування між особинами одного виду.
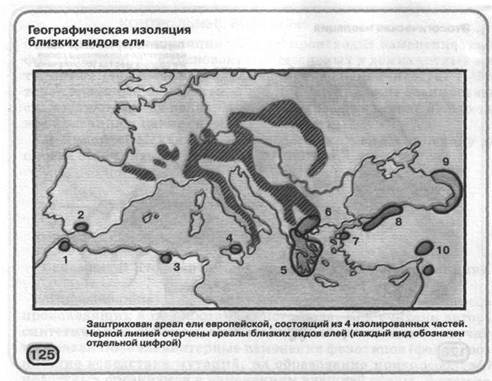
При географічній ізоляції різні популяції одного виду роз'єднані непереборним для них бар'єром (мал. 125). Наприклад, якщо ріка або гірський хребет виникають на території між двома популяціями виду, особини якого не здатні до плавання або польоту; коли лісовий масив розділяється на окремі ліси ділянками із трав'янистою рослинністю.
Екологічна ізоляція спостерігається, коли усередині популяції виникають різні групи особин (екологічні форми, або раси), кожна з яких відрізняється вимогами до умов існування.
Часто утворяться групи, що відрізняються видом їжі. Наприклад, одні раси жука горохової зерновки харчуються насіннями гороху, інших - квасолі. У період розмноження через різні місця виростання кормових рослин особини цих рас між собою не контактують.
Ізоляція може виникати через різні строки розмноження. Наприклад, цвітіння одного виду лугових трав залежить від
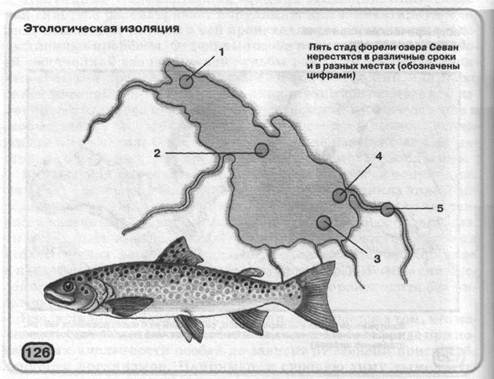
часу припинення весняного паводка: особини, які довго перебували під водою, цвітуть пізніше тих, які взагалі не затоплювалися або попадали під вплив паводка лише на короткий час. Часто ізоляцію забезпечують особливості поводження. Наприклад, в озері Севан (Вірменія) живуть кілька черід форелі, які разом харчуються, однак для розмноження особини кожної череди направляються в особливе місце (мал. 126).
Іноді обмін спадкоємною інформацією можливий лише між певними особинами усередині популяції. Наприклад, в інфузорії-туфельки відомо кілька десятків «підлог», здатних попарно обмінюватися спадкоємною інформацією (конъюгировать) лише в певних комбінаціях. Кожна особина певного «підлоги» виділяє особливу речовину, що діє на поверхневі рецептори клітини іншого «підлоги». Кон'югація можлива тільки при взаємній відповідності цих речовин і рецепторів. Якщо штучно ввести ядро однієї особини усередину інший, несумісної з першої, то злиття генеративних ядер не відбувається й туфелька гине.
Різні форми ізоляції діють незалежно одна від інший і найчастіше одночасно в популяціях певного виду.
Еволюційне значення ізоляції полягає в тім, що при відсутності обміну аллельными генами між популяціями в генофонді кожної з них згодом виникають різні мута-
ции й змінюються частоти зустрічальності аллелей, тому ізольовані популяції адаптуються до зовнішнього середовища незалежно одна від іншої. Ізоляція - це необхідна умова виникнення внутрішньовидової дивергенції (розбіжності ознак). При відсутності ізоляції будь-які зміни, що виникли в одній з популяцій, через послідовні схрещування мігруючих між популяціями особин зрештою поширяться по всьому ареалі виду й дивергенції не відбудеться.
Неважко помітити, що всі висновки синтетичної гіпотези стосуються лише популяцій організмів з перехресним заплідненням. Механізми еволюції партеногенетических і самоплідних організмів, а також видів, у яких полове розмноження відсутнє, до останнього часу практично не вивчені.
Поиск по сайту: