АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
Схрещуванні
" Q /— /^-^ 7\ ЙВ| Ь
\ р\ ':' " в (I '■ | жовтий зелений (I I J
\ Гамети) (?f ^^«^Г _^^
«:: (®^I>tj)®
(62)
Використовуючи мал. 62, простежимо за гомологичными хромосомами при моногибридном схрещуванні гомозиготних особин гороху посівного. Для спрощення допустимо, що певні особини мають тільки одну пару гомологичных хромосом (тобто кількість хромосом у диплоидном наборі рівняється двом: 2п = 2), а кожна із хромосом містить тільки один ген. Хромосома з домінантною аллелью (А) на малюнку позначена жовтим цвітом, а з рецессивной (а) - зеленим. Відомо, що гібриди першого покоління гетерозиготны (їхній генотип - Аа), оскільки одну хромосому з домінантної аллелью вони одержують від одного з батьків, а іншу, з рецессивной, - від іншого. Таким чином, вони единообразны як по генотипі, так і по фенотипі.
У гібридів другого покоління, на відміну від батьківських форм, полові клітини будуть двох сортів: половина з них буде нести хромосому з домінантної аллелью, половина - з рецессивной. У результаті схрещування гібридів першого покоління між собою можливі три варіанти генотипів їхніх нащадків: чверть гібридів другого покоління буде мати хромосоми тільки з домінантними (гомозиготы по домінантної аллели -АА), половина -одну хромосому з домінантної, іншу - з рецессивной (гетеро-зиготы - Аа) і чверть - хромосоми тільки з рецессивными (гомозиготы по рецессивной аллели - аа) аллелями. По фенотипі три чверті насінь гібридів другого покоління (гомозиготы по доми-
нантной аллели й гетерозиготы) будуть мати жовту й одна чверть (гомозиготы по рецессивной аллели) - зелене фарбування.
Таким чином, за умови значної кількості й рівної життєздатності гамет різних сортів стає зрозумілим статистичний характер закону розщеплення, що визначається більшою кількістю равновероятностных зустрічей цих гамет. Результати проведеного Г. Менделем моногибридного схрещування можна записати схематично:

Однак простіше записати хід схрещування за допомогою так званих ґрат Пеннета (названа на честь англ. генетика Р.К. Пен-нета, уперше її що предложили). Ці ґрати зображують так: по горизонталі записують гамети однієї (у раздельнополых організмів - батьківської) особини, а по вертикалі - іншої (материнської). У місцях перетинання горизонтальної й вертикальної ліній указують генотипи нащадків:

Запис схрещування гомозиготних батьківських форм у вигляді ґрат Пеннета наочно демонструє закономірний характер однаковості гібридів першого покоління. Результати ж схрещування між собою гібридів першого покоління будуть такими:

Таким чином, ми бачимо, що серед гібридів другого покоління можливе утворення трьох варіантів генотипу, які за умови повного домінування визначають два варіанти фенотипу. Із цієї таблиці стають зрозумілими й причини подальшого розщеплення ознак при самозапиленні гібридів другого покоління. Гомозиготні особини будуть формувати гамети тільки одного сорту й серед їхніх нащадків явище розщеплення не спостерігається. Гетерозиготні особини будуть формувати два сорти гамет (з домінантної аллелью й з рецессивной), тому серед їхніх нащадків відбудеться розщеплення по фенотипі в співвідношенні 3:1.
Аналогічно можна продемонструвати цитологічні основи й статистичний характер закону незалежного комбінування станів ознак. Представимо, що в диплоидном наборі рослини мають дві пари гомологичных хромосом (2п = 4), кожна з яких несе тільки один ген. Хромосоми однієї пари зобразимо у вигляді паличок, іншої - кружками. Хромосоми, що несуть домінантну аллель, офарбимо в жовтий цвіт, а рецессивную - у зелений (мал. 63).
Нехай материнський організм містить хромосоми тільки з домінантними аллелями, а батьківський - тільки з рецессивными. Такі гомозиготні організми в процесі формування гамет будуть утворювати тільки один їхній сорт - або з домінантними, або з рецессивными аллелями генів фарбування й структури поверхні насіння. При схрещуванні батьківських форм всі особини гібридів першого покоління одержать по двох хромосоми з відповідними домінантними аллелями від материнського організму й рецессивными - від батьківського. Таким чином, всі вони будуть гетерозиготними по двох парах аллелей і будуть формувати чотири сорти гамет у рівних кількостях. При цьому у двох з них аллельные гени будуть перебувати в тих же комбінаціях, що й у гаметах вихідних батьківських форм, а у двох інші -у нових сполученнях (рекомбінаціях). Рекомбінація (від лат. ре -приставка, що позначає повторення дії, і комбинатио -з'єднання) - це з'єднання аллелей різних генів у гаметах гібридних особин, що відрізняється від їхніх комбінацій у гаметах батьків.
Внаслідок рівної ймовірності з'єднання під час схрещування різних сортів гамет гібридів першого покоління, у їхніх нащадків (гібридів другого покоління) можливе утворення дев'яти варіантів генотипів, що визначають чотири різних фенотипи. У двох з них стану ознак цвіту насіння й структури його поверхні сполучаються так само, як і у фенотипі вихідних батьківських форм (жовтий цвіт - гладка поверхня насіння й зелений цвіт - зморшкувата поверхня), а у двох - у нових комбінаціях (жовтий цвіт - зморшкувата поверхня насіння й зелений цвіт - гладка поверхня). Таким чином,
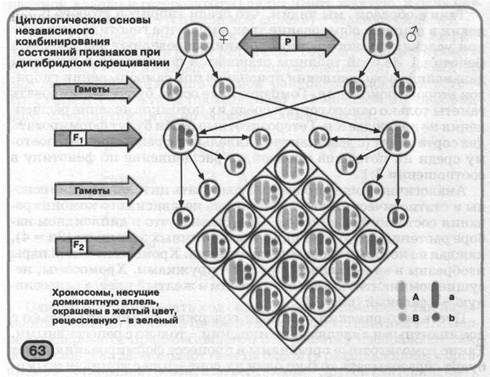
рекомбінації - це одне із джерел комбинативной мінливості.
Статистичний характер закону незалежного комбінування станів ознак можна довести, записавши хід дигибрид-ного схрещування за допомогою ґрат Пеннета. При схрещуванні гомозиготних батьківських форм його результат буде таким (А и а - домінантна й рецессивная аллели гена фарбування, а В и b -відповідно гена структури поверхні насіння):

Таким чином, результати схрещування батьківських форм, гомозиготних по двох домінантним і двох рецессивным аллелям генів фарбування й структури поверхні насіння, розташованих у негомологичных хромосомах, підтверджують закон однаковості гібридів першого покоління: всі вони гетерозиготны по
цим генам і будуть утворювати насіння жовтого цвіту із гладкої
поверхнею.
Хід схрещування гібридів першого покоління між собою
показаний на малюнку 61. Він свідчить, що серед гібридів другого покоління буде спостерігатися таке розщеплення по генотипі: 1 ААВВ: 2 ААВЬ: 1 AAbb: 2 Аавв: 4 Аавь: 2 Aabb: 1 аавв: 2 аавь: 1 aabb, а по фенотипі: 9 частин жовтих насінь із гладкою поверхнею, 3 частини жовтих насінь зі зморшкуватою поверхнею, 3 частини зелених насінь із гладкою поверхнею й 1 частина зелених насінь зі зморшкуватою поверхнею. Таким чином, розщеплення серед гібридів другого покоління як по ознаці фарбування насіння, так і по ознаці структури його поверхні буде становити 12:4 (тобто 3:1). За умови повного домінування домінантних аллелей над відповідними рецессивными, фенотип жовтого насіння із гладкою поверхнею буде визначатися чотирма варіантами генотипу (ААВВ, ААВЬ, Аавв, Аавь), жовтого зі зморшкуватої - двома (AAbb, Aabb), зеленого із гладкої - двома (аавв, аавь) і зеленого зі зморшкуватої - одним (aabb).
Поиск по сайту: