АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
ЯВИЩЕ ЗЧЕПЛЕНОГО СПАДКУВАННЯ. ХРОМОСОМНА ТЕОРІЯ СПАДКОВОСТІ
Згадаєте! Що таке кроссинговер і схрещування, що аналізує?
Явище зчепленого спадкування. Закон незалежного комбінування станів ознак ґрунтується на таких положеннях:
- розвиток різних станів ознак спричиняються ал-лельные гени, що займають однакове положення в гомологичных хромосомах;
- гамети й інші гаплоидные клітини, що мають по однієї гомологичной хромосомі з кожної пари, несуть тільки один аллель-ный ген;
- гени, що контролюють розвиток ознак, наслідуваних незалежно, розташовані в негомологичных хромосомах.
Коли ми доводили статистичний характер законів спадковості, установлених Г. Менделем, то для спрощення допускали, що кожна із хромосом несе тільки один ген. Однак учені вже давно звернули увагу, що кількість спадкоємних ознак організмів значно перевищує число хромосом у гаплоидном наборі. Так, у гаплоидном наборі мухи-дрозофилы, класичного об'єкта генетичних досліджень, є всього чотири хромосоми. Однак кількість спадкоємних ознак і, відповідно, генів, які їх кодують, значно більше. Це означає, що в кожній хромосомі розташовано багато генів. Таким чином, разом з ознаками, наслідуваними незалежно, повинні бути й такі, які успадковуються сцепленно один з одним, оскільки вони визначаються генами, розташованими в одній хромосомі. Такі гени утворять групу зчеплення. Кількість груп зчеплення в організмів певного виду дорівнює кількості хромосом у гаплоидном наборі (напр., у дрозофилы 1 п = 4, у людини 1 п = 2 3).
Експериментально явище зчепленого спадкування довів видатний американський генетик Т. Х. Морган (мал. 1) зі своїми співробітниками.
Слід зазначити, що Т.Х. Морган, так само як у свій час Г. Мендель, вдало вибрав об'єкт для своїх досліджень - муху-дрозофілу. Цих комах легко містити в лабораторіях, вони відрізняються високою плідністю й швидкою зміною поколінь (при оптимальних умовах змісту нове покоління з'являється кожні півтори^-дві тижня), невеликою кількістю хромосом, що значно спрощує спостереження.
Явище зчепленого спадкування Т. Х. Морган установив за допомогою такого експерименту (мал. 66). Він схрестив самців дрозофилы, гомозиготних по домінантним аллелям фарбування тіла (сіра) і форми крил (нормальні), із самками, гомозиготними по відповідної рецессивным аллелям (чорне фарбування тіла - недорозвинені крила). Всі гібриди першого покоління, отримані від такого схрещування, були гетерозиготны по обох генах і мали сіре фарбування тіла й нормальну форму крил. Після цього Т.Х.Морган провів схрещування, що аналізує: він схрестив гібриди першого покоління з особинами, гомозиготними по відповідної рецессивным аллелям.
Теоретично від такого схрещування можна було очікувати два варіанти розщеплення. Якби гени, що визначають фарбування тіла й форму крил, були розташовані в негомологичных хро-о мосомах, тобто успадковувалися незалежно, розщеплення повинне було бути таким: 25% особин із сірим тілом і нормальною формою крил, 25% - із сірим тілом і недорозвиненими крильми,

Слід зазначити, що Т.Х. Морган, так само як у свій час Г. Мендель, вдало вибрав об'єкт для своїх досліджень - муху-дрозофілу. Цих комах легко містити в лабораторіях, вони відрізняються високою плідністю й швидкою зміною поколінь (при оптимальних умовах змісту нове покоління з'являється кожні півтори^-дві тижня), невеликою кількістю хромосом, що значно спрощує спостереження.
Явище зчепленого спадкування Т. Х. Морган установив за допомогою такого експерименту (мал. 66). Він схрестив самців дрозофилы, гомозиготних по домінантним аллелям фарбування тіла (сіра) і форми крил (нормальні), із самками, гомозиготними по відповідної рецессивным аллелям (чорне фарбування тіла - недорозвинені крила). Всі гібриди першого покоління, отримані від такого схрещування, були гетерозиготны по обох генах і мали сіре фарбування тіла й нормальну форму крил. Після цього Т.Х.Морган провів схрещування, що аналізує: він схрестив гібриди першого покоління з особинами, гомозиготними по відповідної рецессивным аллелям.
Теоретично від такого схрещування можна було очікувати два варіанти розщеплення. Якби гени, що визначають фарбування тіла й форму крил, були розташовані в негомологичных хро-о мосомах, тобто успадковувалися незалежно, розщеплення повинне було бути таким: 25% особин із сірим тілом і нормальною формою крил, 25% - із сірим тілом і недорозвиненими крильми,
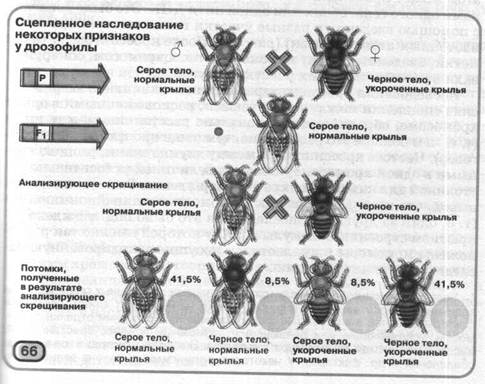
25% - із чорним тілом і нормальними крильми й 25% - із чорним тілом і недорозвиненими крильми (тобто в співвідношенні 1:1:1:1). Якби ці гени розташовувалися в одній хромосомі й успадковувалися сцепленно, то серед нащадків можна було б очікувати 50% особин із сірим тілом і нормальною формою крил і 50% - із чорним тілом і недорозвиненими крильми (тобто в співвідношенні 1:1).
Реально ж 41,5 % нащадків мали сіре тіло й нормальну форму крил, 41,5% - чорне тіло й недорозвинені крила, 8,5% -сіре тіло й недорозвинені крила й 8,5% - чорне тіло й нормальну форму крил. Таким чином, розщеплення по фенотипах наближалося 1:1 (як у випадку зчепленого спадкування). Однак серед нащадків, отриманих у результаті схрещування, що аналізує, були представлені чотири варіанти фенотипів, які можна було очікувати у випадку, якби досліджувані гени успадковувалися незалежно. На підставі цього Т.Х. Морган припустив, що гени, що визначають фарбування тіла дрозофіл і форму їхніх крил, розташовані в одній хромосомі й успадковуються сцепленно, однак при утворенні полових клітин у процесі мейозу гомологичные хромосоми можуть обмінюватися ділянками, тобто спостерігається явище, назване перехрестом хромосом, або кроссинговером, з яким ви познайомилися в § 18.
Кроссинговер підсилює комбинативную мінливість, сприяючи утворенню нових варіантів з'єднань аллелей. При цьому може відбуватися обмін декількома генами або ділянками одного гена, обох або однієї нитки ДНК. Це було доведено за допомогою введення в різні ділянки гомологичных хромосом ізотопних атомів (міток) (мал. 67). Після кроссинговера такі мітки, уведені в одну з гомологичных хромосом, виявляли у відповідних ділянках іншої.
Дослідження кроссинговера виявили такі закономірності: сила зчеплення між двома генами, розташованими в одній хромосомі, обернено пропорційна відстані між ними (тобто чим більше ця відстань, тим частіше відбувається кроссинговер). Частота кроссинговера між двома генами, розташованими в одній хромосомі, є величиною щодо постійної для кожної конкретної пари генів. Був також зроблений висновок, що гени розташовані в хромосомі в лінійному порядку (тобто один за іншим). Згодом це було підтверджено відкриттям будови молекули ДНК, у якої саме так розташовані мономери - нуклеотиды, що несуть закодовану спадкоємну інформацію.
Частота кроссинговера між різними парами зчеплених генів є величиною щодо постійної, однак на неї можуть впливати деякі фактори зовнішнього й внутрішнього середовища (зміни в будові окремих хромосом, що ускладнюють або роблять неможливим процес кроссинговера; висока або низька температура; рентгенівські промені; деякі хімічні сполуки й ін. фактори). У деяких організмів виявлена залежність

частоти кроссинговера від віку (напр., у дрозофилы) або підлоги (напр., у мишей, курей).
Генетичні карти хромосом. Відносна сталість частоти кроссинговера між різними парами генів однієї групи зчеплення використають як показник відстані між окремими генами, а також для визначення послідовності їхнього розташування в хромосомі. Вивчення частот кроссинговера між різними парами генів уможливило створення генетичних карт хромосом (мал. 68), на яких показаний порядок розташування й відносні відстані між генами в певній хромосомі. Для деяких організмів (напр., кукурудзи, декількох видів дрозофіл) уже створені повні генетичні карти всіх хромосом, для інших (напр., людина) -тільки частково.
Хромосомная теорія спадковості. Результати досліджень Т.Х. Моргана і його співробітників лягли в основу запропонованої ними хромосомної теорії спадковості, що багато в чому визначила подальший розвиток не тільки генетики, але й біології в цілому. Вона дозволила з'ясувати матеріальну основу законів спадковості, установлених Г. Менделем, і встановити, чому в певних випадках спадкування ознак від них відхиляється. Основні положення хромосомної теорії спадковості такі:

- гени розташовані в хромосомах; кожна з негомологичных хромосом має свій унікальний набір генів;
- гени розташовані в хромосомах у лінійному порядку;
- кожний ген займає в хромосомі певна ділянка; ал-лельные гени займають однакові ділянки в гомологичных хромосомах;
- всі гени однієї хромосоми утворять групу зчеплення, завдяки чому відбувається зчеплене спадкування деяких ознак; сила зчеплення між двома генами, розташованими в одній хромосомі, обернено пропорційна відстані між ними;
- зчеплення між генами порушується в результаті обміну ділянками гомологичных хромосом (кроссинговера) у профазі першого мейотического розподілу;
- кожний біологічний вид характеризується певним хромосомним набором (каріотипом) - кількістю й особливостями будови окремих хромосом.
Поиск по сайту: