АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомДругоеЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция
СИСТЕМИ СХРЕЩУВАНЬ ОРГАНІЗМІВ І ЇХНІ ГЕНЕТИЧНІ НАСЛІДКИ
Згадаєте! Що таке летальні й су б летальні аллели, гомозигота й гетерозигота?
Гібридизація і її форми Ефективність селекції залежить не тільки від форми штучного добору, але й від правильного підбора батьківських пар виробників і застосування тієї або іншої системи схрещування організмів - гібридизації.
Гібридизація - процес одержання гібридів, заснований на об'єднанні генетичного матеріалу різних клітин або організмів. Гібриди утворяться в результаті полового процесу або шляхом об'єднання нестатевих клітин. В останньому випадку ядра таких гібридних клітин можуть зливатися з утворенням загального ядра або ж залишатися відособленими. Гібридизація можлива як у межах одного виду (внутрішньовидова), так і між особинами різних видів або навіть пологів (міжвидова, або віддалена). Внутрішньовидове схрещування буває близкородственным і неспорідненим.
Близкородственное схрещування - це схрещування організмів, що мають безпосередніх загальних предків. Залежно від ступеня генетичної спільності близкородственное схрещування може бути більш-менш тісним. Найбільш тісні форми близкородственного схрещування спостерігаються серед самопильних рослин і гермафродитних тварин, для яких характерне самозапліднення. В організмів з перехресним заплідненням найбільш тісні форми близкородственного схрещування спостерігаються при спарюванні братів із сестрами, батьків з їхніми нащадками.
Внаслідок близкородственного схрещування з кожним наступним поколінням гібридів підвищується їх гомозигот-ность. Це пояснюється тим, що чим більше генетична подібність батьківських форм, тим вище ймовірність з'єднання в генотипі нащадків тих самих аллелей різних генів. У самопильних рослин уже в 10-м поколінні спостерігається майже повна гомозиготность (до 99,9%), а при схрещуванні братів із сестрами або батьків з нащадками подібний результат може бути досягнуть після 20-го покоління. Однак 100% гомозиготне ти по всіх генах досягти не вдається, оскільки вона порушується виникаючими мутаціями.
Біологічним наслідком близкородственного схрещування є ослаблення або навіть виродження нащадків (мал. 77). Це пояснюється тим, що в нащадків, отриманих у результаті близкородственного схрещування, підвищується ймовірність переходу в гомозиготний стан рецессивных летальних або сублетальных аллелей, у результаті чого вони можуть виявитися у фенотипі.
Таким чином, тісне близкородственное схрещування часто приводить до появи організмів з різними спадкоємними аномаліями. Наслідку близкородственного схрещування були відомі людині здавна. Наприклад, приблизно 20% людей-альбіносів є нащадками від близкородственных шлюбів. У людини відомо трохи рецессивных летальних аллелей, здатних у гомозиготному стані привести до смерті. Тому шлюби між близькими родичами в багатьох народів уважалися небажаними або ж взагалі заборонялися релігією або законами.
У селекції близкородственное схрещування використають для одержання чистих ліній. Воно дозволяє перевести в гомозиготний стан аллели, що визначають коштовні для селекціонерів стану ознак.
Неспоріднене схрещування. Явище гетерозису. Неспоріднене схрещування - гібридизація організмів, що не мають тісних родинних зв'язків, тобто представників різних ліній, сортів або порід одного виду. Неспорідненими вважають особин, у яких загальні предки відсутні як мінімум протягом останніх шести поколінь. Неспоріднене схрещування застосовують для з'єднання в нащадках коштовних якостей, властивим представникам різних ліній, порід або сортів. По своїх генетичних наслідках неспоріднене схрещування прямо

протилежно близкородственному. При неспорідненому схрещуванні з кожним наступним поколінням підвищується гетерозиготне ть нащадків. Це пояснюється тим, що зі зменшенням ступеня споріднення організмів зростає ймовірність наявності в них різних аллелей певних генів.
У випадку неспорідненого схрещування часто спостерігається явище гетерозису, або «гібридної сили» (мал. 78). Гетерозис (від греч. гетероиозис - зміна, перевтілення) - явище, при якому перше покоління гібридів, отримане в результаті неспорідненого схрещування, має підвищені життєздатність і продуктивність у порівнянні з вихідними батьківськими формами. Це пояснюється тим, що в гетерозисных форм сублетальные й летальні рецессивные аллели переходять у гетерозиготний стан і їхній несприятливий вплив не проявляється у фенотипі. Крім того, у генотипі гібридних особин можуть з'єднуватися сприятливі домінантні аллели обох батьків. Внаслідок цього може відбуватися взаємодія домінантних аллелей неаллельных генів.
Найбільше чітко гетерозис проявляється в першому поколінні гібридів. У наступних же поколіннях, внаслідок розщеплення ознак і переходу частини генів у гомозиготний стан, ефект гетерозису слабшає й до восьмого покоління сходить на немає. У рослин ефект гетерозису можна закріпити

вегетативним розмноженням, подвоєнням числа хромосом або партеногенезом. Гетерозис може більше позначитися на одних ознаках гібридної особини, не зачіпаючи інших.
Явище гетерозису широко застосовують у сільському господарстві, оскільки воно дозволяє значно підвищити продуктивність (напр., у кукурудзи - на 20-25%). Ефект гетерозису добре виражений в овочевих культур (цибулі, томатів, огірків, баклажанов, буряка й ін.). У тваринництві схрещування між різними породами прискорює ріст і полове дозрівання нащадків, поліпшує якість м'яса, молока й т.д.
Так, у результаті схрещування яйценоских порід курей між собою (напр., леггорнів з австралорнами) продуктивність гібридів зросла на 20-25 яєць у рік. У бройлерів (гібридні курчата м'ясних порід) гетерозис проявляється в прискоренні росту й поліпшенні якості м'яса.
Віддалена гібридизація. Подолання стерильності міжвидових гібридів. Перспективним методом селекційної роботи є віддалена гібридизація - схрещування особин, що належать до різних видів і навіть родів з метою з'єднання в генотипі гібридних нащадків коштовних спадкоємних ознак. За допомогою віддаленої гібридизації створені гібриди пшениці й пирію, що відрізняються високою продуктивністю (до 300-450 ц/га зеленої маси) і стійкістю до полягання; пшениці й жита й ін. гібриди. Відомі міжвидові гібриди й серед плодово-ягідних культур (малини й ожини, зливи й терну й т.д.).
У тваринництві також виведені численні міжвидові гібриди. Добре відомий гібрид коня й осла - мул, що відрізняється значною силою, витривалістю й більшою тривалістю життя в порівнянні з батьківськими видами. Подібними властивостями характеризується й гібрид одногорбого й двогорбого верблюдів. Гібрид білуги й стерляді - бестер - швидко росте й має високі смакові якості м'яса.
Однак селекціонери часто зіштовхуються із проблемою безплідності міжвидових гібридів, гамети яких звичайно не дозрівають. Навіть за умови однакової кількості хромосом у кари-отипах батьківських форм, їхні хромосоми відрізняються розмірами й особливостями будови й тому не здатні конъюгиро-вать у процесі мейозу. Особливо ускладнюється хід мейозу за умови різної кількості хромосом у каріотипі батьків.
Якщо в селекції рослин стерильність міжвидових гібридів ще можна перебороти, то в селекції тварин вирішити цю проблему значно складніше. Тільки в окремих випадках у міжвидових гібридів тварин один або обидві підлоги є плідними. Так, у гібрида яка (свійська тварина високогірних районів Центральної Азії) з великою рогатою худобою самці марні, тоді як самки - плідні; а мули взагалі не здатні розмножуватися.
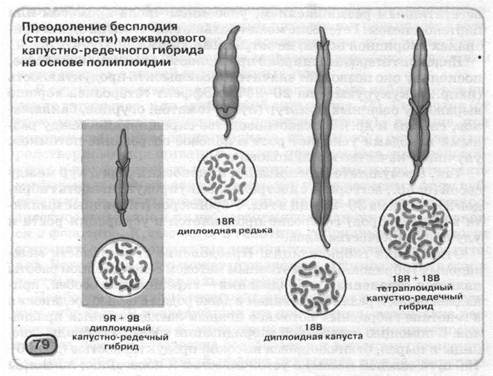
Уперше методику подолання безплідності міжвидових гібридів у рослин розробив в 1924 році Г. Д. Карпеченко на прикладі гібрида капусти й редьки. Цей гібрид по своєму фенотипі займав проміжне положення між відповідними фенотипами батьківських форм (мал. 79). Хоча капуста й редька -представники різних пологів сімейства Капустяні (Крестоцветные), кількість хромосом у них однакове (2п=18). Однак створений Г.Д. Карпеченко гібрид виявився марним, оскільки в процесі мейозу «капустяні» і «редькові» хромосоми не конъюгировали між собою. Тоді вчений подвоїв кількість хромосом гібрида (4п = 36). Таким чином, у ядрах нестатевих клітин гібридів було по двох повних набоору хромосом батьківських форм. У результаті процес мейозу в такий тетраплоидной форми проходив нормально: «капустяні» хромосоми кожної пари конъюгировали з «капустяними», а «редькові» - з «редьковими». Таким чином, у кожну з гамет завжди попадало по одному гаплоидному наборі хромосом редьки й капусти.
Поиск по сайту: